
Нюансы женского тренинга

Эксперт в области бодибилдинга и пауэрлифтинга. Тренируюсь более 20 лет, подписывайтесь на мои соц. сети.
Многие женщины, составляя план тренировок, не учитывают гендерные особенности, работают в спортзале до седьмого пота по мужским программам и удивляются отсутствию должного результата. Из-за разницы строения тела и протекающих в организме физиологических процессов тренировочные программы для мужчин и женщин существенно различаются. Для достижения максимального эффекта от спортивных занятий построение тренировок важно проводить в соответствии с женской физиологией.
Гормоны

У женщин значительно меньше, чем у мужчин, вырабатывается тестостерон — гормон, отвечающий за увеличение мышечной массы. Даже активно тренируясь, представительницы прекрасного пола не смогут быстро нарастить мышцы и не должны опасаться чрезмерной мышечной гипертрофии. Делая базовые упражнения, используя гантели и постепенно увеличивая вес снаряда, женщины подтянут мышцы, создадут рельеф и добьются идеальной фигуры, оставшись при этом женственными: появление перекачанных мускулов им не грозит.
Общий показатель нормы тестостерона для мужчин колеблется от 345 до 950 нг/дл, у женщин показатели в разы ниже — от 15 до 70 нг/дл.
Уровень женских возможностей в силовых тренировках ниже мужских. Из-за сниженного количества норадреналина женщины не способны тренироваться до мышечного отказа при выполнении упражнения. Они всегда заканчивают подход на несколько повторений раньше.
На образование жировой ткани и ее распределение по женскому телу оказывают влияние эстрогены, синтезируемые в организме в достаточном количестве. Эффект от тренировок достигается только в случае гормонального баланса. Как недостаточное, так и избыточное количество женских половых гормонов ведет к появлению лишнего веса вне зависимости от физических нагрузок.
Циклические особенности

Здоровый женский организм работает как часы, в соответствии со строгим циклическим режимом. Для достижения максимальных результатов от тренировок важно опираться на менструальный цикл при распределении тренировочной нагрузки.
- Активные силовые тренировки целесообразно проводить в первые 2 недели цикла. Фолликулярная фаза — оптимальное время для повышения нагрузок и уровня сложности занятий. В этот период женщины наиболее выносливы, легче переносят болевые ощущения, а жировые отложения благодаря быстрому сжиганию углеводов тают на глазах.
- Наивысшая мышечная сила и предел физических возможностей наблюдаются во время овуляции. Максимально эффективно в эти дни расходуются питательные вещества. Овуляторная фаза подходит для пиковых спортивных нагрузок (в случае, если не планируется беременность).
- В лютеиновую фазу женский организм переключается в режим накопления энергии. Постепенно возрастает утомляемость, повышается риск травмирования, интенсивность тренировок идет на спад. Упражнения на пресс и прокачка ножных мышц становятся малоэффективны. Акцент смещается на кардиотренировки. Если силы отсутствуют, не стоит слишком усердствовать: нужно тренироваться в меру своих возможностей.
- Менструация — время, когда организм не справляется с чрезмерными нагрузками. Можно устроить себе небольшую передышку. При нежелании прекращать спортивные занятия в эти дни предпочтение лучше отдать мягкому фитнесу, йоге и бассейну.
Исследование немецких ученых доказало, что снижение интенсивности тренировок во время лютеиновой фазы помогает женщинам в фолликулярной фазе после возобновления привычных занятий увеличивать выносливость более чем на 30%.
Принцип прогрессии нагрузок

Суть прогрессии нагрузок состоит в том, что от раза к разу тренировки понемногу становятся сложнее: постепенно увеличиваются веса, растет количество повторений или подходов, возрастает интенсивность. Это помогает организму оставаться в тонусе.
Частая женская ошибка — страх перетрудиться в зале, боязнь физической нагрузки. Подобрав оптимальную со своей точки зрения интенсивность спортивных занятий, дамы фиксируют ее и повторяют от тренировки к тренировке. Отсутствие изменений приводит к адаптации организма, привыканию мышц и отсутствию прогресса.
Основной принцип успешного тренинга: нагрузки должны расти. Для результата важно двигаться вперед.
Большое количество повторений

Женщины отличаются от мужчин тем, что способны делать больше повторений в каждом подходе, дольше не чувствуют усталости и быстрее восстанавливаются. Так заложено генетически.
В женских мышцах низок процент миофибрилл, ответственных за сокращение и расслабление мышечного волокна. Если мужчины хорошо набирают мышечную массу за счет работы с большими весами при малом количестве повторов, у женщин этот номер не пройдет. Множество подходов и повторений в женской тренировке обеспечат активное сжигание жира и постепенный рост мышц. Минимальные перерывы между подходами усилят эффект.
Оптимальное количество повторений в подходах — от 12 до 20.
Какие должны быть веса

Женщины менее сильные, чем мужчины, это факт. Поэтому они работают с малыми весами, чтобы предотвратить травмы и не надорваться.
Чтобы правильно подобрать рабочий вес в упражнении, женщина должна ориентироваться на свое самочувствие, желаемую нагрузку и количество выполняемых повторений:
- Новичкам, желающим похудеть, для начала можно выбрать гантели весом до 2 кг;
- Подтянуть мышцы и слегка нарастить мышечную массу помогут снаряды весом 3-10 кг, в зависимости от уровня подготовки;
- Постепенно рабочие веса будут расти, подготовленная женщина в состоянии приседать с весом 30-50 кг.
Не нужно сразу начинать тренироваться с большими весами. Для начального этапа важно отточить технику правильного выполнения упражнений, а затем уже постепенно увеличивать вес утяжеления.
Распределение мышц

Общее соотношение мышц к массе тела у мужчин и женщин не одинаково. Усредненный показатель соотношения мышечной массы к общему весу у женщин составляет около 36%. У мужчин цифры выше — от 40 до 50%.
Распределение мышечной массы по телу также различается. У женщин большее количество мышц сосредоточено в нижней части тела. Акцент в упражнениях делается на ноги и ягодицы, которые прокачиваются быстрее мышц спины, груди и рук. Мышцы верха натренировать гораздо труднее, во время тренировок им требуется уделять больше времени и внимания.
Научный эксперимент показал: если сравнить женщину и мужчину примерно равного уровня натренированности, мышцы верхней части тела женщины окажутся на 40% слабее, мышцы нижней части тела — всего на 20%.
Особую сложность составляет для слабого пола работа с мышцами нижнего пресса. В нижней части живота у женщины мало нервных окончаний, благодаря чему уменьшается чувствительность к менструальным болям, но и реакция данной области на нагрузки также снижается.
Питание

Наравне с физическими нагрузками важную роль в построении красивого тела играет правильная организация питания.
Если главной целью является похудение, важно тщательно следить за калорийностью рациона.
Чтобы организм справлялся с интенсивными физическими нагрузками, с пищей должны в достаточных количествах поступать витамины и минеральные элементы.
Нельзя полностью отказываться от жиров. Правильные жиры положительно влияют на гормональную и сердечно-сосудистую системы женщин. Женщины, которые потребляют больше жира, на тренировках сжигают больше калорий, жмут лежа больший вес, более подтянуты.
Углеводов в питании должно быть немного. Лучше немного недобрать дневную норму, чем превысить ее. Важно свести к минимуму потребление быстрых углеводов: сладостей, выпечки. Именно быстрые углеводы легче всего превращаются в жир, с которым трудно бороться.
Строительным материалом для мышц является белок. Его в пище должно быть достаточное количество, особенно у любительниц силовых тренировок.
Сбалансированное разнообразное питание — залог хорошей фигуры.
Обмен веществ

Обмен веществ в женском организме происходит медленнее, чем в мужском, энергозатраты на каждый килограмм веса для поддержания нормального протекания процессов жизнедеятельности у женщин ниже.
Генетически женщины предрасположены к накоплению жира для нормального функционирования организма и здоровья репродуктивной системы.
Нормальный процент жира в организме женщины составляет от 16 до 25%. Для мужчины достаточно всего 3%.
Мужчины сжигают жир даже в состоянии покоя, а посттренировочное жиросжигание у них длится дольше.
В состоянии покоя женский организм практически не сжигает жиры, питаясь полученной из углеводов глюкозой. Лишь усердные тренировки помогают трансформировать лишние жировые запасы в энергию.
Гликоген в организме

Особенностью женского организма является быстрая переработка калорий в нужное количество гликогена, и такая же быстрая скорость его расходования. Избыточные углеводы быстрее чем у представителей сильного пола трансформируются в жир на боках.
Вместе с тем, женская потребность в гликогене меньше, чем мужская. Организм обычно использует в качестве тренировочного топлива именно подкожный жир.
В результате выполняя одинаковые по интенсивности упражнения женщина сжигает больше жиров, меньше углеводов и белка, чем мужчина.
Основная цель женских тренировок — похудение и коррекция проблемных зон. Нарастить мышцы стремится редкая представительница слабого пола. Адаптация тренировочного процесса под особенности женского организма позволит повысить эффективность занятий, приобрести фигуру мечты без вреда для женского здоровья.
Sektascience: научно-популярный журнал
Пишем статьи о питании, тренировках и здоровом образе жизни на основе научных исследований
Тренировочный план в рамках менструального цикла
Автор: Ольга Реброва, сертифицированный тренер по пилатесу
Совсем недавно мы рассказали, можно ли заниматься пилатесом во время менструации. Но на самом деле очень важно знать, что происходит с женским организмом во время всего цикла, а не только в первые его дни.
Учет фаз менструального цикла дает тренеру и тренирующейся дополнительную подсказку, как распределить нагрузки, чтобы они способствовали развитию физических качеств.

На вопрос о том, стоит ли вообще заниматься спортом в период менструации, можно ответить однозначно: тренироваться стоит, если позволяет самочувствие. Многолетние исследования ученых говорят, что в зависимости от характера протекания менструальной функции, спортсменок можно разделить на 4 группы:
- Первая группа (примерно 50% спортсменок).
Имеют хорошее самочувствие и общее состояние по показателям функциональных проб. Демонстрируют высокие спортивные результаты во время всех фаз цикла. - Вторая группа (примерно треть спортсменок).
В период менструальной фазы испытывают общую слабость, быструю утомляемость, сонливость, отсутствие желания тренироваться. Объективно: наблюдается снижение артериального давления, удлинение восстановительного периода после функциональных проб. - Третья группа (около 5% женщин).
В первые дни цикла испытывают повышенную раздражительность, чувство “скованности”, боль внизу живота, головную боль, беспокойный сон. Объективно: учащается ЧСС и повышается артериальное давление, особенно максимальное. - Четвертая группа (около 5% женщин).
Во время менструации у таких спортсменок развивается комплекс симптомов, подобный проявлениям интоксикации: общее недомогание, тошнота, ноющие боли в суставах, мышцах, беспокойный сон. Объективно: ЧСС и дыхание учащаются, артериальное давление либо в норме, либо понижается.
Для каждой группы предусмотрены свои рекомендации по нагрузке в период менструаций.

Тренировочный план в рамках цикла
| Менструальный цикл (лат. menstruus — месячный, ежемесячный) — это закономерные циклические изменения в организме женщины, повторяющиеся через определённые промежутки времени и внешне проявляющиеся менструацией. |
Ежемесячный менструальный цикл очень индивидуален, но в среднем составляет 21—35 дней. Этот период делится на 5 фаз в зависимости от изменения уровня основных гормонов, которые оказывают влияние на организм. Ниже приведены средние значения продолжительности фаз на примере 28-дневного цикла, у каждой женщины эти значения индивидуальны.
Менструальная фаза (1 – 4 (7) день)
Физиология фазы:
- рост фолликулов и начало активной выработки эстрогена;
- сокращается количество эритроцитов в крови (примерно на 1 млн) и снижается уровень гемоглобина (на 15 %). Такие изменения отмечаются у 50% женщин. На 8-10-й день после первого дня менструации эти показатели возвращаются в норму.
- снижаются аэробные возможности организма;
- при нагрузке повышается частота сердечных сокращений и дыхания;
- могут снижаться мышечная сила, быстрота и выносливость, но улучшается гибкость;
- повышается болевой порог и склонность к травматизму.
Рекомендации
В этот период не стоит ставить в план сложные силовые тренировки и давать акцентированную нагрузку на мышцы брюшного пресса. Если самочувствие в эти дни ухудшается, можно сосредоточиться на мягких практиках, растяжке, расслаблении.
 Низкоинтенсивная и умеренная нагрузка, по мнению специалистов, поможет безболезненно и легко пережить менструацию, снять спазмы в области живота за счет увеличения кровообращения в малом тазу. Низкоинтенсивная и умеренная нагрузка, по мнению специалистов, поможет безболезненно и легко пережить менструацию, снять спазмы в области живота за счет увеличения кровообращения в малом тазу. |
Замените свои привычные тренировки на малоинтенсивные силовые нагрузки (меньшее число подходов), мягкую йогу, пилатес, плавание, пешие прогулки и спокойную езду на велосипеде.
| Поводом для отказа от физической нагрузки в эти дни могут стать болезненные ощущения, общее плохое самочувствие и в целом нежелание заниматься. |
Постменструальная фаза (5(8) – 12(13) день)
Физиология фазы:
- уровень выработки эстрогена резко повышается, стимулируя анаболический эффект в мышечных тканях;
- растет общий уровень прогестерона и тестостерона;
- повышается общая работоспособность и выносливость;
- лучше работает сердечно-сосудистая система.
Рекомендации
Это лучшие дни, чтобы тренировать свою силу и выносливость. Вы и сами, скорее всего, замечали, как после менструаций появляется много сил и энергии. Это прекрасное время для фитнеса, работы с весами над крупными мышцами всего тела, а также для кардионагрузок любого типа.
| Можно заниматься ВИИТ, кроссфитом, сайклингом, кикбоксингом и проходить всевозможные интенсивные программы по типу bootcamp. |
Овуляторная фаза (13 –15 день)
Физиология фазы:
- уровень эстрогена только начинает снижаться с самой высокой отметки, а уровень прогестерона еще невелик;
- происходит снижение работоспособности, физическая работа требует от организма больше затрат;
- женщина может ощущать слабые боли внизу живота, повышение аппетита и либидо;
- у женщины увеличивается отдел мозга, отвечающий за эмоции и воспоминания (исследование учёных из Института Макса Планка);
- женщина находится на пике своей фертильности.
Рекомендации
Высокий уровень эстрогена негативно влияет на метаболизм коллагена, поэтому риск получить травму в этот период достаточно высокий.

Профессиональные тренеры рекомендуют сбавить обороты и заняться йогой, танцами, балетом, пилатесом, можно устроить себе длинные кардиосессии — это обычный бег в среднем темпе на свежем воздухе или бег на эллиптическом тренажере в зале.
Постовуляторная фаза (16 – 24 день)
Физиология фазы:
- эстроген начинает снижаться, а прогестерон же, напротив, начинает активно повышаться;
- возможны перепады настроения, повышение температуры тела до 1 градуса;
- снижение работоспособности, ощущение вздутия и отечность;
- обмен веществ становится менее активным по сравнению с фолликулярной фазой, и организм не так охотно сжигает жир.
Рекомендации
Снизить неприятные ощущения в теле лучше всего поможет сбалансированное рациональное питание.
Для физически подготовленных девушек в эту фазу цикла рекомендованы тренировки, которые помогают организму использовать жир в качестве топлива. Сил на данном этапе меньше, поэтому подойдут короткие интенсивные занятия, недолгий интервальный бег на кардиотренажере, интенсивная аэробика, силовые или активные виды йоги (аштанга-виньяса-йога), тай-бо, плавание.
Для менее подготовленных рекомендуются умеренные нагрузки, групповые силовые классы в фитнес-клубе, хатха-йога, силовой пилатес, плавание и занятия на фитболе.
| Тренировки дадут естественный всплеск эндорфинов, поднимут настроение, заставят чувствовать себя лучше, а усиленное в этот период потоотделение при физической нагрузке поможет вывести лишнюю жидкость из организма. |
Предменструальная фаза (25 – 28 день)
Физиология фазы:
- концентрация эстрогена и прогестерона в крови падает, снижая функциональные возможности организма;
- вероятно подавленное настроение, раздражительность, апатия;
- возможен незначительный набор веса (это нормально!).
Рекомендации
Тренеры разрешают пропустить пару занятий в конце цикла, если состояние совсем не располагает к выполнению упражнений. Силовая нагрузка должна быть умеренной.
| По сравнению с другими фазами цикла, интенсивность занятий в этот период должна быть наименьшей. |

Снять неприятные симптомы предменструальной фазы помогут пешие прогулки на свежем воздухе и плавание.
Исследователи из Университета спорта в Ванкувере утверждают, что женщины, которые регулярно занимаются фитнесом, сообщают о меньшей болезненности груди, более редких вздутиях живота и перепадах настроения в сравнении с нетренирующимися.
| Если ваш цикл больше или меньше 28 дней, стоит внимательно понаблюдать за собой в течение 2-3 месяцев, чтобы более точно определить моменты перехода из одной фазы в другую. |
Влияние фаз цикла на физическое и психологическое состояние
Менструальная функция начинается в период полового созревания и продолжается до 45-55 лет. Регулярный цикл устанавливается сразу после менархе (период, когда менструация появилась впервые) или в течение последующих 1,5 – 2 лет.
Продолжительность его исчисляется от первого дня наступившей менструации до первого дня следующей менструации и характеризуется довольно широким диапазоном индивидуальных физиологических колебаний (от 21 до 35 дней, в среднем — 28).
Нормальный цикл состоит из двух основных фаз изменения функций яичников: фолликулярной и лютеиновой.
- Фолликулярная фаза (может длиться от 11 до 27 дней в зависимости от индивидуального цикла). За это время происходит формирование доминирующего фолликула, и наступает овуляция (выход яйцеклетки из фолликула).
- Лютеиновая фаза (длится от 11 до 17 дней). После выхода яйцеклетки из фолликула в яичнике образуется желтое тело, которое выделяет эстроген и прогестерон для подготовки организма к беременности.
На физиологическое состояние, желание и возможность женщины посвящать себя физической активности влияют уровень гормонов и внешние факторы.
- Изменение уровня гормонов в разных фазах цикла.
Наибольшее влияние на женский организм во время менструального цикла оказывают эстрогены (эстрадиол), гестагены (прогестерон) и андрогены (более всего тестостерон). - Внешние факторы.
За последние десятилетия набралось достаточное количество наблюдений, которые показывают, что тренировки на профессиональном уровне или регулярные интенсивные занятия кроссфитом могут привести к нарушениям репродуктивной системы спортсменок.
Исследования и мнения
Основные исследования в этой области касаются женщин-спортсменок и влияния их биологического цикла на спортивные результаты. Известно, что меньше всего течение менструального цикла сказывается на женщинах-спринтерах и больше всего – на спортсменках, тренирующихся на выносливость.
Согласно многочисленным исследованиям в различных спортивных дисциплинах, самыми неблагоприятными фазами цикла для серьезных физический нагрузок являются фазы 1, 3 и 5. В эти фазы регистрировались самые низкие показатели скоростных, силовых, скоростно-силовых упражнений, а также скоростной выносливости.
| Непосредственно в дни менструации не рекомендуется выполнять силовые упражнения, которые сопровождаются натуживанием, резкими движениями и охлаждением тела. Объем силовых нагрузок в эти дни должен быть небольшим, важнее всего ориентироваться на самочувствие. |

Установлено, что с увеличением стажа спортивных выступлений и ростом спортивного мастерства заметно снижается отрицательное влияние так называемых неблагоприятных фаз на специальную работоспособность (работоспособность, связанная с определенным видом спорта). Таким образом, чем женщина тренированней, тем меньшее негативное влияние фаз 1,3 и 5 она испытывает, при условии отсутствия проблем с менструальным циклом из-за серьезных физических нагрузок.
Оптимизация тренировки в зависимости от дня цикла
В 2014 году было проведено исследование, целью которого было определить изменение силовых способностей и психо-эмоционального состояния женщин в зависимости от фаз МЦ. Под наблюдением находилось 12 женщин в возрасте 20-25 лет. Исследование включало анкетирование и практические тесты.
В анкете выясняли субъективное отношение участницы эксперимента к вопросу, её состояние и реакции организма на физические нагрузки, наличие изменений в протекании цикла. Исследователей интересовало, связывают ли спортсменки эти изменения с проведенными тренировками, что предпринимают в случае сбоев цикла. Все женщины отметили связь специфики тренировок с характером протекания цикла.
Наибольший объем тренировок всегда давали в те периоды, когда организм женщин предрасположен к ее выполнению. Спортсменкам, у которых 28-дневный цикл, увеличивали физические нагрузки на 7 – 12 и 16 – 25 дни, снижали на 1 – 6 и 13 – 15 дни цикла.
В ходе всего цикла применялись упражнения избирательного характера по группам мышц. Например, за 1–2 дня до и в дни менструаций выбирались упражнения, не вызывающие большого напряжения мышц нижней части брюшного пресса и малого таза.
Выявлено, что динамика проявления специальной выносливости и скоростных возможностей имеет подъемы и спады соответственно фазам менструального цикла.
Наиболее высокие показатели проявились у девушек в начале постменструальной и постовуляторной фаз цикла, что соответствует пикам содержания половых гормонов и связанных с ними колебаний работоспособности спортсменок.
На практике ваш тренер или вы сами должны учитывать снижение работоспособности и психофизиологических функций организма в разные фазы цикла.
Максимально усложнять программу тренировок оптимально в постменструальные и постовуляторные дни. В период сниженной работоспособности необходимо избегать тяжелых физических нагрузок и чаще отдыхать.
Выводы
Менструальный цикл — это автоматизированный, налаженный, задуманный природой процесс. Это точно не болезнь, а лишь особенность женского организма, но особенность важная, поскольку отвечает за репродуктивную функцию.
Физическая активность, которая учитывает состояние женщины в каждый момент времени, способствует безболезненному и спокойному течению цикла.
Зная закономерности и цикличность распределения гормональных изменений, а также наблюдая за собой в течение месяца, вы можете самостоятельно корректировать свой тренировочный график. В рамках одного направления можно увеличивать и уменьшать интенсивность занятий. В рамках разных — планировать так, чтобы более интенсивные тренировки совпадали с фазами повышения работоспособности.
При этом важно помнить, что всё индивидуально, нужно обязательно прислушиваться к своему организму. Тренируйтесь на благо своего здоровья и не забывайте отдыхать, когда этого требует ваше тело.
Адаптация мышц к силовой тренировке
Адаптация мышц к силовой тренировке мышц проявляется в виде морфологической адаптации: увеличения площади поперечного сечения целой мышцы и отдельных мышечных волокон, происходящее вследствие увеличения размера и числа миофибрилл, гиперплазии миофибрилл, изменений в типе волокна, архитектуре мышц, миофибриллярной плотности, а также в структуре соединительной ткани и сухожилий и неврологической адаптации: обучения и координации в работе мышц, переносе обучения с одной конечности на другую.
Folland J.P., Williams A.G. The Adaptations to Strength Training Morphological and Neurological Contributions to Increased Strength// Sports Med. 2007; 37 (2): 145-168
Фолланд Дж., Вильямс А.
АДАПТАЦИЯ К СИЛОВОЙ ТРЕНИРОВКЕ. МОРФОЛОГИЧЕСКИЙ И НЕЙРОФИЗИОЛОГИЧЕСКИЙ ВКЛАД В УВЕЛИЧЕНИЕ СИЛЫ
РЕЗЮМЕ
Гипертрофическая силовая тренировка (HRST) – одна из самых широко практикуемых форм физической активности, используемая для увеличения атлетических показателей, улучшения опорно-двигательного аппарата и повышения эстетики тела. Регулярная активность этого типа обеспечивает заметный прирост в мышечной силе, который ассоциируется с рядом неврологических и морфологических адаптаций. В настоящем обзоре оцениваются факты, свидетельствующие об этих адаптациях, их взаимодействии, их вкладе в увеличение силы, а также вопросы методологии исследования.
Первичные морфологические адаптации включают увеличение площади поперечного сечения целой мышцы и отдельных мышечных волокон, происходящее вследствие увеличения размера и числа миофибрилл. Клетки-сателлиты активируются на очень ранних стадиях тренировки; их быстрое увеличение и последующее слияние с существующими волокнами тесно связано с гипертрофией мышцы. Другие возможные морфологические адаптации включают гиперплазию мышечных волокон, изменения в типе волокна, архитектуре мышц, миофибриллярной плотности, а также в структуре соединительной ткани и сухожилий.
Косвенные свидетельства неврологических адаптаций, включающих обучение и координацию, возникают из-за специфичности тренировочной адаптации, переноса обучения одной конечности на контралатеральную конечность и предполагаемым сокращениям мышцы. Видимое увеличение напряжения, специфичного для целой мышцы, в первую очередь использовалось для доказательства неврологических адаптаций; однако морфологические факторы (например, предпочтительная гипертрофия мышечных волокон (МВ) второго типа, увеличенный угол перистости МВ, увеличение радиологической плотности) также с большой вероятностью вносят свой вклад в этот феномен. Критически важными выглядят изменения внутримышечной координации. Адаптация в активации мышц-агонистов, как показывает электромиография, тетаническая стимуляция и техника вызванной стимуляции, предполагают малые, но существенные изменения. Увеличение частоты возбуждающих нервных импульсов и спинальные рефлексы являются наиболее вероятными причинами этого улучшения, хотя имеются факты, предполагающие отсутствие изменений в корковой или корково-спинальной возбудимости.
Увеличение силы при HRST, несомненно, объясняется широкой комбинацией неврологических и морфологических факторов. Хотя неврологические факторы могут внести самый большой вклад на ранних стадиях тренировочной программы, гипертрофические процессы также начинаются в начале тренировок.
ВВЕДЕНИЕ
Гипертрофическая силовая тренировка (HRST) – один из наиболее распространённых видов физической активности. В первые недели тренировок с отягощениями сила значительно вырастает, и прирост длится порядка 12 месяцев [1]. Такой тип тренировки направлен на улучшение спортивных результатов, оздоровление опорно-двигательного аппарата и повышение эстетики тела. Несмотря на то, что гипертрофическая силовая тренировка чаще всего ассоциируется с атлетическими видами спорта, требующими большой силы и мощи, она может быть использована и при заболеваниях или их последствиях (саркопения, нарушения опорно-двигательного аппарата, иммобилизация, повреждения или долгий постельный режим), она также положительно влияет на метаболические процессы в мышце. Также HRST хорошо развивает выносливость [2]. Таким образом, HRST вызывает значительный научный интерес. В данной статье оценены доказательства этих адаптаций, их взаимодействие и вклад в увеличение силы, а также применяемые методики.
1. МОРФОЛОГИЧЕСКИЕ АДАПТАЦИОННЫЕ ИЗМЕНЕНИЯ
1.1. ИЗМЕНЕНИЯ РАЗМЕРОВ МЫШЦЫ
Несложно заметить, что у лиц, тренирующихся с отягощениями, через несколько месяцев наблюдается видимый прирост мышечной массы. Это было широко описано в научной литературе. Исследования с использованием целого ряда методов сканирования (например, магнитно-резонансной томографии [МРТ]; компьютерной томографии [КТ], и ультразвука), как правило, обнаруживали значительное увеличение анатомического поперечника (площади поперечного сечения мышцы) в течение относительно короткого периода тренировок (8-12 недель) [3-6]. МРТ считается лучшим методом определения площади поперечного сечения мышцы из-за большого разрешения [7], и используется преимущественно в последнее десятилетие. В кропотливом и более длительном исследовании (Narici et al.[8]) рассмотрены изменения мышечной силы, площади поперечного сечения мышцы (МРТ) и активность мышц-агонистов (посредством электромиографии) за шесть месяцев стандартных тренировок с отягощениями (рис. 1). Они продемонстрировали, что рост объема мышцы (гипертрофия), вначале изменяется по линейному закону. Даже после шести месяцев тренировки плато не появляется. Однако после двух месяцев тренировки сила четырехглавой мышцы бедра и ее анатомический поперечник (ACSA) перестают развиваться параллельно. Интуитивно понятно, что рост скелетной мускулатуры должен быть более медленным и иметь плато. Количественные доказательства представили Always et al. [9] на бодибилдерах, имеющих стаж тренировок более пяти лет. Они не нашли изменений в анатомическом поперечнике двуглавой мышцы плеча после 24 недель тренировки.
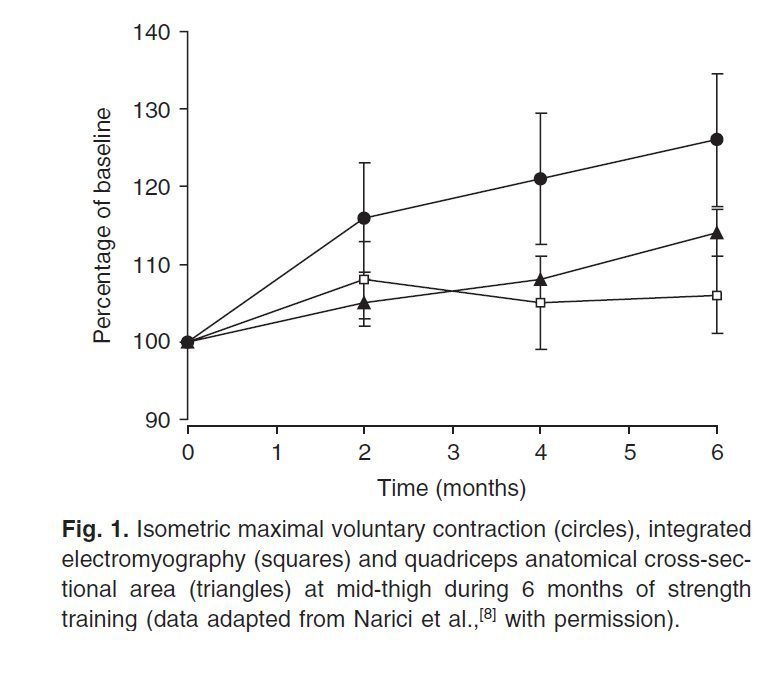
Рис.1. Изометрическое максимальное сокращение (кружки), интегрированная ЭМГ (квадраты) и ACSA четырехглавой мышцы бедра измеренный на середине бедра (треугольники) после 6 месяцев силовой тренировки (адаптированная данные Narici et al. [8] с разрешения авторов).
Другое общее наблюдение, связанное с HRST — непропорциональное увеличение силы мышц по сравнению с ACSA, что указывает на увеличение напряжения всей мышцы. Существуют многочисленные методологические проблемы, связанные с прямым сравнением этих параметров, главным образом методология измерения размера мышц. В подавляющем большинстве исследований измеряется ACSA (анатомический поперечник – площадь поперечного сечения мышцы в самой утолщенной части), то есть всего на одном уровне. И этот показатель является как индексом размера мышц. Недавнее исследование надежности измерения размера мышц пришло к заключению, что измерение площади поперечного сечения (CSA) всего на одном уровне менее надежно, чем многократное измерение этого показателя в различных сечениях. Оно должно использоваться только там, где ожидается относительно большое изменение в размере мышцы [10]. Теоретически, физиологический поперечник (PCSA), измеренный перпендикулярно к ходу мышечных волокон, казался бы более точным индексом сократительной способности мышцы. Однако, точное измерение PCSA проблематично [11], так как требуется измерение объема мышц и угла перистости волокна, а также оценка длины волокна [12]. Альтернативно, в некоторых исследованиях оцениваются изменения в объеме мышц посредством МРТ после гипертрофической тренировки (+14%, за 12 недель тренировки мышц-сгибателей предплечья [13]; +9.1%, за 12 недель тренировки межостных мышц позвоночника [14]; +12%, за 9 недель тренировки четырехглавой мышцы бедра [5]; +10%, за 14 недель тренировки четырехглавой мышцы бедра [15]). Вопрос о том, что размеры мышц являются самый точным индикатором мышечной силы, оспаривается. Bamman et al. [16] пришел к заключению, что ACSA и PCSA более сильно коррелированы с работой силы; однако, Fukanaga et al. [17] нашел более высокие корреляции, между PCSA, а также объемом мышц с максимальным моментом силы, чем у ACSA. Еще одним фактором является то, что измерения размера мышц под влиянием HRST до настоящего времени проводились только в пассивном состоянии мышцы. Даже во время изометрического сокращения, контрактильные элементы сокращаются и могут внести значительные изменения в морфологию мышц и механику опорно-двигательного аппарата [18,19]. Например, у медиальной головки икроножной мышцы от состояния расслабления до максимального произвольного сокращения в неподвижном (изометрическом) положении, происходят следующие изменения: угол перистости мышечного волокна удваивается, а PCSA увеличивается на 35% [20]. Различные индексы, характеризующие размер мышц (ACSA, PCSA или объем мышц), как показывают измерения на МРТ, существенно изменяются после 8–12 недель регулярной тренировки. Эта адаптация, кажется, продолжается в линейном виде в течение первых шести месяцев тренировки. К сожалению, самый точный показатель размера мышц до сих пор не найден, и до сих пор не обращается внимания на проблему смешивания измерений размеров мышц, проведенных в покое.
1.1.1. Влияние группы мышц
У ранее нетренированных людей гипертрофия мышц наблюдается больше в мышцах верхних конечностей по сравнению с нижними [21,22]. При стандартной тренировке Welle et al. [23] обнаружили, что анатомический поперечник (ACSA) мышц-сгибателей предплечья увеличился на 22% и 9%, у молодых и пожилых мужчин соответственно, в то время как у мышц-разгибателей голени он увеличился соответственно лишь на 4% и 6%. Последние исследования плотности мышц (оценивается с помощью ультразвука) показали более высокую реакцию на стандартные упражнения для верхних мышц тела по сравнению с мышцами нижних конечностей [6]. Возможным объяснением этого является тот факт, что мышцы нижних конечностей, в частности, антигравитационные мышцы: четырехглавая бедра и трицепс голени, обычно всегда активны и испытывают более значительные нагрузки во время повседневной жизнедеятельности, чем верхние мышцы тела [22] и таким образом, меньше реагируют на данный раздражитель нагрузки. Альтернативным объяснением этого, являются межмышечные различия в содержании андрогенных рецепторов. При этом есть некоторые доказательства большей концентрации этих рецепторов в мышцах верхней части тела по сравнению с мышцами нижних конечностей [24].
1.1.2. Влияние пола
В среднем, скелетные мышцы женщины обладают 60-80% силы, площадью поперечного сечения мышечного волокна и анатомическим поперечником (ACSA) мышцы мужчины [25-28]. Таким образом, не удивительно, что абсолютные изменения в силе и объеме мышц после тренировки у женщин меньше [22] в соответствии с их меньшими размерами [29]. Более низкий уровень андрогенов в крови у женщин также вызывает для нижней части тела меньшую относительную гипертрофию мышц на тренировке, по сравнению с мужчинами [30-32].
Для нижних конечностей в ряде исследований не удалось найти никаких различий между мужчинами и женщинами, как в плане гипертрофии, так и в силе после гипертрофической силовой тренировки [6, 22, 33-37]. Например, Tracy et al. [5] сравнивая гипертрофию четырехглавой мышцы у пожилых мужчин и женщин после девяти недель гипертрофических силовых тренировок в результате показал 12% увеличение объема мышц у обеих групп. Наоборот, результаты с тренировкой верхней части тела указывают, что при гипертрофических силовых тренировках могут быть различия между полами [38-40].
Последнее масштабное исследование 342 женщин и 243 мужчин показало, что после 12 недель идентичных тренировок, у мужчин наблюдается большее увеличение анатомического поперечника (+2,5%, МРТ), а у женщин – большее увеличение силы (+25%, при использовании 1-ПМ; +6% в изометрических сокращениях) [39]. Потенциально большая гипертрофия мышц верхней части тела мужчин может быть связана с большей концентрацией андрогенных рецепторов в этих мышцах [24], что и заставляет их более чутко реагировать на более высокое содержание в крови андрогенов.
Большая сила мышц у женщин, возможно, отражает большую способность к нейрогенной адаптации [41]. Она, возможно, могла быть намного больше у мышц верхней части тела, но из-за меньшего воздействия на них в повседневной жизни, этого не происходит.
1.1.3. Влияние возраста
Нет никаких сомнений, что пожилые люди, в том числе люди старческого возраста (от 90 до 99 лет), подвергаются гипертрофии скелетных мускулов в ответ на HRST (ACSA середины бедра возрастает на +9% после 8 недель тренировки [42] и на +9,8% после 12 недель тренировки [43]). Абсолютный прирост мышечной массы меньше в старческом возрасте по сравнению с людьми пожилого возраста, вероятно, из-за меньшего размера мышц типичного для старческого возраста [23]. Некоторые сравнительные исследования показывают, что относительное изменение объема мышц или ACSA (анатомического поперечника) в ответ на HRST не зависит от возраста [34, 44], тогда как другие предлагают меньший гипертрофический ответ у пожилых людей [14, 23, 45]. Вариативность результатов, скорее всего, обусловлена низкой численностью испытуемых, принимающих участие в этих исследованиях и большой межиндивидуальной изменчивостью в ответ на HRST [39].
1.1.5. Избирательная гипертрофия
При увеличении объема всей мышцы было обнаружено варьирование в росте ее головок, а также длины каждой из головок мышц [4, 8, 46, 47].
Например, Housh et al. [4] сообщили, что объем прямой мышцы бедра в среднем увеличился на 23,2%, в то время как у латеральной широкой бедра только на 7,5% (рис. 2). Narici et al. [8] нашел, что гипертрофия прямой мышцы бедра варьирует в пределах от 10% до 50% в различных местах вдоль мышцы.
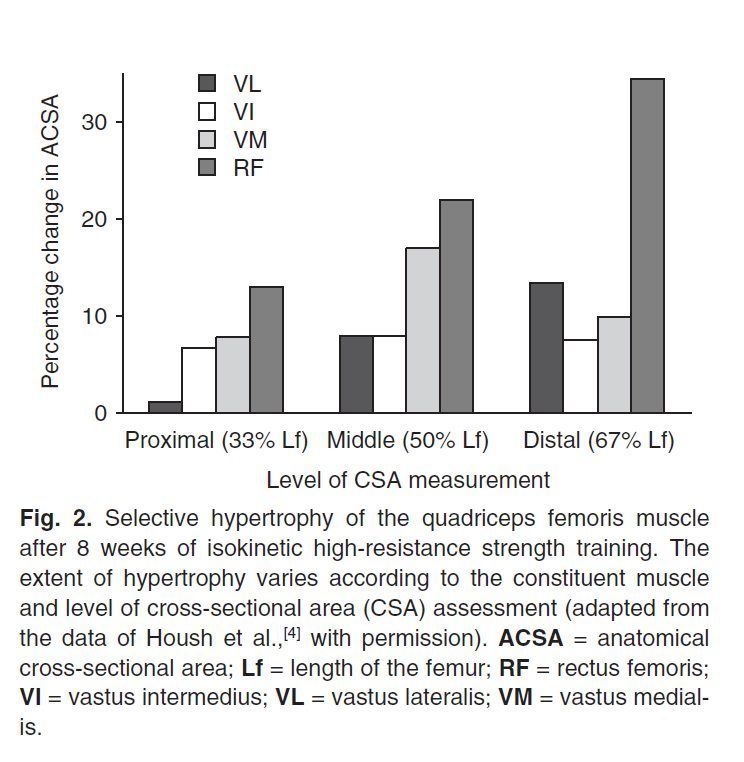
Рис.2. Селективная (избирательная) гипертрофия четырехглавой мышцы бедра после 8 недель изокинетической гипертрофической тренировки. Степень гипертрофии изменяется в соответствии с составом мышц и площадью поперечного сечения (CSA). (Рисунок адаптирован на основе данных Housh et al. [4].)
Эти авторы предположили, что гипертрофия каждого компонента мышцы может сильно зависеть от степени их нагрузки (напряженности) и активации, которые, вероятно будут регулироваться механикой каждой составляющей мышц под воздействием тренировок.
Например, четыре головки четырехглавой мышцы бедра могут иметь разные соотношения «длина-напряжение» и, следовательно, различный вклад во вращающий момент (момент силы) под любым углом при совместной работе.
Некоторые исследования показали наибольший ответ гипертрофической реакции всей четырехглавой мышцы и двуглавой мышцы плеча, в области максимального обхвата (площади поперечного сечения), например, обхват середины бедра [5, 13, 48].
Однако другие исследователи нашли, что наибольшая гипертрофическая реакция происходит только в проксимальных [46] или проксимальных и дистальных [8] отделах мышц. Возможно, это связано с различиями в выполняемых упражнениях.
Имеются свидетельства, что это явление селективного (избирательного) роста может продолжаться в течение длительного периода времени.
Для проведения исследований были взяты юноши-тяжелоатлеты (средний возраст 16,4 года). После 18 месяцев тренировки, анатомический поперечник (ACSA) четырехглавой мышцы бедра увеличился на 31% на расстоянии 30% длины бедра, считая от колена (Lf), но он не изменился на расстоянии 50% Lf или 70% Lf [49].
Для определения точного количественного показателя роста мышечной ткани в перспективе необходимо сканировать несколько срезов посредством МРТ.
Теоретически, рост мышечной массы, может быть достигнут либо путем увеличения площади поперечного сечения мышечных волокон (гипертрофия волокна), увеличения числа волокон (гиперплазии волокна) или увеличения длины волокон, которые первоначально не соответствовали длине мышцы.
1.2. ГИПЕРТРОФИЯ МЫШЕЧНОГО ВОЛОКНА
Увеличение площади поперечного сечения мышечных волокон (мышечная гипертрофия) обычно расценивается, как первичная адаптация к долгосрочной силовой тренировке и была широко документирована (см. обзоры McDonagh and Davies [50] и Jones et al. [51]). Гипертрофия мышечных волокон, как полагают, вносит вклад в увеличение площади поперечного сечения мышцы, способствует увеличению сократительного материала (количеству поперечных мостиков) расположенных параллельно и, таким образом, увеличивает силовые показатели мышцы. Изменения в площади поперечного сечения мышечного волокна можно оценить, на основе биопсии скелетных мышц. Имеются сообщения о широком варьировании изменений площади поперечного сечения мышечного волокна в ответ на гипертрофическую силовую тренировку (HRST). Тренировка трехглавой мышцы плеча в течение шести месяцев привела к гипертрофии волокон I и II типа на 27 и 33% соответственно [52]. Aagaard et al. [11] нашли, что после 14 недель силовых тренировок с сопротивлением (resistance training) увеличение в среднем площади поперечного сечения мышечных волокон составляет 16%. Это увеличение достоверно коррелирует с увеличением объёма мышцы. В то время как подавляющее большинство исследований показали значительное увеличение площади поперечного сечения мышечного волокна, Narici и соавторы [8] не обнаружили никаких изменений в средней площади мышечного волокна, несмотря на то, что анатомический поперечник мышцы (максимальная площадь поперечного сечения мышцы) увеличился на 19%. Такое варьирование результатов может быть обусловлено рядом факторов, в том числе: (1) плохой воспроизводимостью техники биопсии; (2) индивидуальной реакцией человека на тренировку, и (3), направленностью тренировки (различные режимы работы мышц, тип и скорость сокращений, интенсивность и продолжительность работы). Была установлена низкая надежность (повторяемость) измерений площади поперечного сечения мышечного волокна с одним образцом биопсии (коэффициент вариации был равен 10-24%) [53-57]. Это происходит, из-за большой неоднородности размеров мышечных волокон в пределах скелетной мышцы, которая зависит от глубины места биопсии [57], а также изменчивости перпендикулярных срезов мышечной ткани и установления границ мышечного волокна [56]. Таким образом, в то время как масса доказательств решительно поддерживают гипертрофию мышечного волокна, к данным от одиночных образцов биопсии следует относиться с осторожностью [59].
1.2.1. Преимущественная гипертрофия мышечных волокон II типа
После силовой тренировки обнаруживают преимущественную гипертрофию мышечных волокон II типа [60-63]. Данные, представленные Hakkinen et al. [64] показывают большие пластические возможности мышечных волокон II типа. Обычно они более быстро увеличиваются в размерах во время тренировок и быстрее атрофируются без тренировочного процесса.
Поэтому неудивительно, что многие кратковременные исследования (6-10 недель) показывают гипертрофию только у мышечных волокон II типа [11, 63, 65, 66], тогда как длительные исследования показывают увеличение площади поперечного сечения волокон как I типа, так и II типа [52-54]. Исследования на животных доказывают, что мышечные волокна II типа проявляют большую гипертрофическую реакцию на нагрузку [67]. Процентное соотношение в мышце мышечных волокон II типа достоверно коррелирует с гипертрофией, вызванной тренировкой [45] и возрастанием силы [65]. Тем не менее, прирост силы не связан с композицией мышечных волокон [68] и имеет положительную связь с соотношением мышечных волокон I типа.
Это говорит о том, что волокна II типа осуществляют большее напряжение, что приводит к преимущественной гипертрофии этого типа волокон. Их преимущественное увеличение способствует повышению силы мышцы, что часто наблюдают во всей мышце во время тренировки. Однако по поводу напряжения различных типов мышечных волокон было много дискуссий. Fitts et al. [69], заключили, что нет существенных различий в напряжении различных типов мышечных волокон у человека или крысы. Наоборот, более современные труды говорят о большем напряжении мышечных волокон у людей, имеющих изоформу тяжелого миозина второго типа (MHC IIX), по сравнению с волокнами, имеющими МНС I типа (+50% [70], +20% [71], +32% [72]). Исследования, которые связывают изометрическое напряжение и композицию волокон, полученные на живом человеке, противоречат полученным данным [73-75]. Однако, процент содержания в мышце волокон II типа (или содержание MHCII) положительно коррелирует с изокинетической силой на скоростях от средних до высоких [76] и относительной силой на высоких [73, 77].
Современные данные указывают на то, что мышечные волокна II типа производят значительно большее напряжение, что, в сочетании с гипертрофической реакцией, вероятно, способствует увеличению в целом напряжения мышцы.
1.3. МИОФИБРИЛЛЯРНЫЙ РОСТ И ПРОЛИФЕРАЦИЯ
Постоянство плотности упаковки миозиновых филаментов внутри миофибрилл указывает на то, что миофибриллярный рост осуществляется посредством добавления сократительных белков на периферии миофибрилл. Более того, исследования показали, что новообразовавшиеся белки склонны образовываться вокруг периферии существующих миофибрилл [79]. Увеличение поперечного сечения миофибрилл, несомненно, способствует увеличению поперечного сечения мышечного волокна. Однако, непропорционально большое увеличение площади поперечного сечения мышечного волокна (вдвое больше, чем площадь поперечного сечения миофибрилл) предполагает дополнительную адаптацию. Постоянство плотности упаковки миозиновых филаментов и увеличивающееся число «расщепленных» миофибрилл после тренировки, теория MacDougal et al. [52] интерпретирована как доказательство увеличения числа миофибрилл после тренировки (т.е. пролиферацию).
1.3.1. Возможный механизм миофибриллярной пролиферации
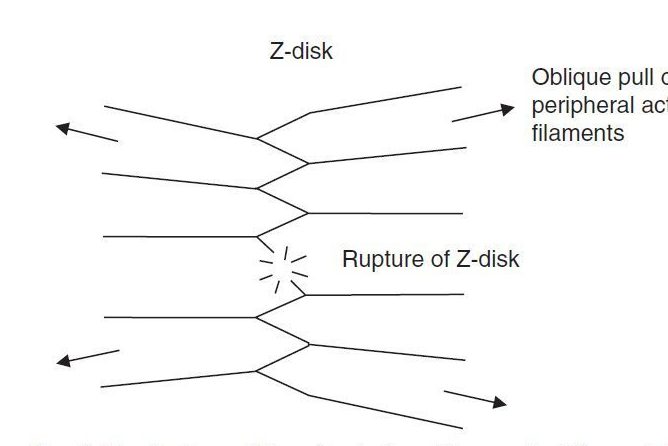
Исследования MacDougal et al. [52,78] показали, что рост и пролиферация миофибрилл являются основными морфологическими изменениями, в ответ на работу, вызывающую рост мышц в организме человека. Во время нормального роста мышц млекопитающих, было обнаружено 15-ти кратное увеличение числа миофибрилл. В серии исследований роста мышей после рождения, Goldspink [80,81] и Howells [82] предложили механизм миофибриллярной пролиферации. Расхождение в массивах, формирующих диски А и I в саркомере вызывает то, что актиновые филаменты растягиваются под несколько косым углом относительно Z-дисков. По мере увеличения размера миофибрилл, периферийные филаменты будет подвергаться большим боковым смещением относительно А- и Z-дисков, и будут тянуть под определенным углом (рис. 3). Goldspink [80,81] предположил, что если это было бы достаточно распространено в обоих полусаркомерах, это могло бы привести к разрыву Z-диска.
Рис. 3. Разрыв миофибрилл в месте прикрепления актиновых филаментов (перерисовано с разрешения автора Goldspink [83])
Как только один Z-диск разрывается, следующий Z-диск может разрушиться таким же образом, пока вся миофибрилла не разделится вдоль своей длины. Доказательства расщепления миофибрилл и разрыва Z-диска приводящие к пролиферации миофибрилл, были обнаружены при росте мышечной ткани птиц и рыб. Таким образом, в ответ на рост, а также, вероятно в ответ на HRST, пролиферация миофибрилл происходит в результате разрывов Z-дисков и продольного деления, которое ограничивает размер миофибрилл и способствует их эффективному контролю и регулированию.
1.3.2. Клетки — сателлиты
Многие исследования выявили, что соотношение количества ядер к объему саркоплазмы остается везде практически неизменным на фоне широкого спектра условий роста (у животных [86,87] и у людей [88,89]). В мышцах человека, Landing et al. [90] выявили прямую корреляцию между количеством ядер и диаметром мышечного волокна. Следовательно, получается, что одно миоядро в состоянии поддержать определенный объем саркоплазматического материала, и это соотношение, по-видимому, будет примерно в два раза больше для мышечных волокон II типа, по сравнению с мышечными волокнами I типа [89].
Работа с животными показала, что во время нормального роста и созревания, увеличение размера мышечного волокна происходит из-за прибавления новых ядер, возникающих из клеток-сателлитов [86,87] . В отличие от ядра, расположенного внутри мышечного волокна (миоядра), клетка-сателлит, находящаяся под базальной мембраной, которая окружает каждое волокно, может подвергаться митозу (делению) и, как правило, одна из дочерних клеток затем становится ядром [91]. Новые миоядра, полученные из клеток-сателлитов, так как они более не способны делиться, начинают производить определенные мышечные белки, которые увеличивают размер мышечного волокна [92,93].
В мышцах взрослых кошек, которые были подвергнуты нагрузке отягощением, Allen et al. [94] обнаружили, что увеличение числа миоядер больше, чем соответствующее увеличение объема мышечного волокна. Rosenblatt et al. [95-97] изучили изменения в скелетных мышцах млекопитающих в ответ на нагрузку по модели, в которой удаляется синергист. Эти авторы сообщили о значительно меньшей гипертрофии после предварительного облучения мышц, которое предотвращает деление клеток-сателлитов. На основе этого они пришли к выводу, что быстрое увеличение количества клеток-сателлитов является необходимым условием для гипертрофии мышцы, следующей за удалением мышц-синергистов.
В исследованиях на человеке Kadi et al. [98,99] показал, что число клеток-сателлитов и число миоядер выше у элитных бодибилдеров по сравнению с контрольной группой (число ядер больше на 35% в мышечных волокнах I типа, и на 31% больше в мышечных волокнах II типа) [98]. Эти авторы сделали вывод о том, что экстремальная гипертрофия мышечных волокон этих атлетов зависит от повышенного содержания миоядер. Продолжительные исследования гипертрофической силовой тренировки продемонстрировали увеличение (прирост) популяции клеток-сателлитов через 9-14 недель тренировок [100-102]. Последние исследования показывают быстрое увеличение (пролиферацию) количества клеток-сателлитов в течение четырех дней после однократной тренировки с использованием эксцентрических упражнений [103]. Однако, влияние гипертрофической силовой тренировки на число миоядер и на отношение числа ядер к объему саркоплазмы спорное. В ответ на 10 недель постоянных тренировок, Kadi и Thornell [100] сообщили, что число миоядер и клеток-сателлитов в трапециевидной мышце увеличилось в значительно большей степени, чем площадь поперечного сечения мышечных волокон (рис. 4).
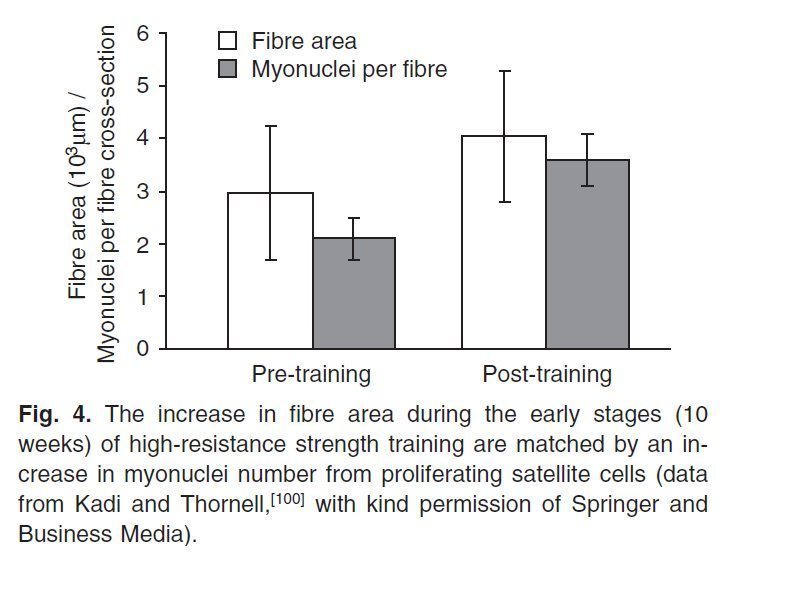
Рис. 4. Возрастание поперечного сечения мышечного волокна на первой стадии (10 недель) силовой тренировки (high-resistance strength) и количества миоядер из-за пролиферации клеток-сателлитов (данные Kadi и Thornell [100]).
Они пришли к выводу, что появление дополнительных миоядер, было необходимо для утолщения мышечных волокон, которое следует даже после краткосрочных силовых тренировок. Hikida et al. [104] также обнаружили, что соотношение числа миоядер и объема саркоплазмы осталось неизменным после 16 недель силовых тренировок, что привело к 30% увеличению размера мышечных волокон. Тем не менее, Kadi et al. [102] сообщили об отсутствии изменений числа миоядер и об увеличении области мышечного волокна, контролируемом каждым миоядром через 90 дней гипертрофической силовой тренировки. Рассматриваемые в целом, эти результаты исследований показывают, что первоначально гипертрофия вызывает ограниченное увеличение количества миоядер и количества саркоплазматических белков, поддерживаемых каждым ядром, но впоследствии потребуется дополнительные миоядра, полученные из клеток-сателлитов.
Для того чтобы возникла гипертрофия, должны быть произведены дополнительные сократительные белки и они должны быть функционально интегрированы в существующие мышечные волокна и миофибриллы. Этот чистый прирост мышечных белков явно требует постоянного избытка синтеза над распадом. Повышенный синтез белка зависит от транскрипции или трансляции и его описание выходит за рамки настоящего обзора. Обзоp «Новые миоядра, полученные из клеток-сателлитов, так как они более не способны делиться, начинают производить определенные регуляции синтеза белка» был сделан Sartorelli и Fulco [105].
1.4 ГИПЕРПЛАЗИЯ
Гиперплазия – увеличение количества мышечных волокон, вызванное продольным расщеплением существующего мышечного волокна [106] с последующей гипертрофией дочернего волокна и/или посредством миогенеза [107]. Любой из этих процессов может способствовать увеличению поперечного сечения мышцы и возрастанию силы в ответ на гипертрофическую силовую тренировку. Однако как явление гиперплазия остается спорной.
1.4.1 Исследования на животных
Индуцированное расщепление мышечных волокон посредством работы наблюдается у животных, что приводит к гиперплазии [108-110]. Методология изучения мышечной гиперплазии – подсчет количества мышечных волокон только на одном уровне мышцы, ставит все получаемые результаты под вопросом. Даже в мышцах с параллельным ходом мышечных волокон не все волокна могут иметь одно и то же место начала и прикрепления. Поэтому в некоторых исследованиях использовалась техника азотной кислоты для расслоения и прямого подсчета клеток-сателлитов [86,87] и общего количества волокон. С помощью подсчета общего количества мышечных волокон Gollnik et al. [111] искал ответ на гипертрофию мышц, которая происходила при удалении агониста и растяжения мышцы (опыты проводились на крысах). Они не нашли никаких доказательств гиперплазии и сделали вывод, что увеличение объема мышцы происходит за счет гипертрофии существующих мышечных волокон. Gonyea et al. [112] подсчитывали количество волокон у кошек после приблизительно 101 недели силовых тренировок. Было обнаружено достоверное увеличение количества волокон (9%; p<0,05), которое было связано с новообразованием их из клеток-сателлитов, но не было представлено доказательств продольного расщепления мышечных волокон.
Обзор 17 исследований Kelly [113] показал, что в мышцах млекопитающих гиперплазия выражена слабее по сравнению с мышцами птиц (соответственно 8% против 21% для мышцы птиц). При подсчете количества волокон посредством техники азотной кислоты гиперплазия наблюдается в 11% случаев, в то время как при использовании гистологического подсчета – в 21%. Таким образом, степень гиперплазии, зависит от экспериментального протокола. Например, в исследованиях с перегрузкой или растягиванием вызывается гиперплазия, в то время как при использовании упражнений гиперплазия или значительно меньше или вообще отсутствует.
1.4.2. Исследования на человеке
Этические и методологические проблемы подсчета количества мышечных волокон у человека в естественных условиях, делают проведение исследований гиперплазии у людей крайне сложной задачей. Даже при исследованиях на трупах существуют большие межиндивидуальные различия, которые связаны с различной адаптацией человека [115]. Пролиферативная способность скелетной мышечной ткани при регенерации хорошо документирована [116]. Appel et al. [117] обнаружили свидетельства формирования новых миотуб (мышечных трубочек) из клеток-сателлитов через шесть недель тренировки на выносливость. В ответ гипертрофическую силовую тренировку Кади и Thornell [100] обнаружили миотубы, а также маленькие мышечные волокна, экспрессирующие эмбриональные и неонатальные тяжелые цепи изоформ миозина. Тем не менее, Appel [107] предположил, что из-за медленных темпов формирования новых волокон, гиперплазия оказывает очень незначительный эффект на поперечное сечение мышцы и следовательно ее силы. Исследования трупов Sjostrom et al. [115] поддержали идею гиперплазии у взрослых людей, однако эти изменения проявляются очень медленно. Чтобы опровергнуть возможность гиперплазии, было проведено сравнение среднего размера мышечных волокон у тренированных (гипертрофическая силовая тренировка) и нетренированных людей [54,118-121]. С учетом ранее обсуждавшейся вариативности средней площади мышечного волокна, полученной посредством биопсии; способствовать увеличению площади поперечного сечения мышцы и возрастанию силы в ответ на гипертрофическую силовую тренировку. Однако как явление, часто в сочетании с низким количеством исследуемых, это может привести к ошибочным выводам. Несколько более достоверным является определение количества волокон путем деления площади поперечного сечения мышцы, полученной посредством компьютерной томографии или МРТ на среднюю площадь волокна, измеренную посредством биопсии. Это зависит от экстраполяции постоянной площади волокна и угла перистости на всю мышцу, как правило, на основе одного образца биопсии [111], который, как говорилось в разделе 1.2, может быть не репрезентативен для измерений площади поперечного сечения мышечного волокна. Используя эту технику, Always et al. [122] сообщили данные о достоверной корреляции между количеством мышечных волокон и анатомическим поперечником у элитных бодибилдеров, которые могут выражать либо адаптивную реакцию на силовую тренировку или процесс отбора. В ответ на трехмесячную гипертрофическую силовую тренировку, MacCall et al. [123] не обнаружили никаких изменений в количестве мышечных волокон, хотя площадь поперечного сечения мышцы возросла на 10%. Сравнение количества мышечных волокон у элитных бодибилдеров и нетренирующихся мужчин не обнаружило достоверных различий [124].
Количественный вклад гиперплазии в изменения площади поперечного сечения мышц человека в ответ на упражнения во многом остается неизвестным. Тем не менее, исследования человека и млекопитающих мышцы предлагают, что гиперплазия составляет, в лучшем случае, небольшую часть увеличения площади поперечного сечения мышцы в ответ на повышенную нагрузку.
1.5. ДРУГИЕ МОРФОЛОГИЧЕСКИЕ АДАПТАЦИИ
1.5.1. Изменения в типе мышечных волокон и композиции тяжелых цепей миозина?
Большинство исследований по мышечной адаптации к силовой тренировке представляют доказательства, свидетельствующие против существенных изменений типа мышечных волокон. У животных количественные методы, используемые для изучения роста мышц не обнаружили никаких изменений в типе волокон при гипертрофии или атрофии [67,125,126], хотя последние исследования показали, что более тонкие изменения могут произойти, а именно переход мышечных волокон типа IIB в тип IIX [127]. У людей тренировка с сопротивлением (resistance training) также, кажется, влияет на изменения типа мышечных волокон. Некоторые исследования показали, достоверное увеличение числа волокон типа IIА и сопутствующим падением количества мышечных волокон типа IIX волокна, [45,60,61,128]. В одном исследовании показано, что эти изменения происходят после 18 тренировочных занятий [129].
Системы классификации мышечных волокон, применяемые в настоящее время, базируются на выделении MHC изоформ. Schiaffino et al. [130] выделены четыре отдельных MHC изоформ (I, IIA, IIB, IIX). Большинство волокон можно отнести к одному типу MHC изоформы, что отражает их функциональные и метаболические свойства, и в целом соответствует другим системам классификации мышечных волокон. Показано, что доля MHC IIX уменьшается на 5-11% с аналогичным ростом МНС IIA после 12-14 недель тренировки. [131-133]. Williamson et al. [132] исследовали композицию мышечных волокон до и после 12 недель гипертрофической тренировки. Этими авторами найдено увеличение доли волокон МНС IIA (+24% у молодых женщин и +27% у молодых мужчин) за счет сокращения доли гибридных волокон (MHC I/IIА и IIA/IIX). В заключение следует заметить, что тонкие изменения типов мышечных волокон и композиции мышечных волокон, по всей видимости, происходят на ранней стадии (2-3 месяца) тренировки, но нет никаких доказательств того, что эти изменения продолжается в течение длительного периода.
1.5.2 Плотность скелетных мышц и миофиламентов
Общая радиологическая плотность скелетных мышц увеличивается после силовых тренировок на +3%; [134] и на +5% [135, 136]. Sipila and Suominen [137] найдено 11% увеличение радиологической плотности трехглавой мышцы голени после 18 недель силовых тренировок у пожилых женщин. Это измерение плотности включает в себя гораздо более широкие слои мышечной ткани, чем исследования плотности упаковки нитей миозина, рассмотренных MacDougall et al. [52] и включает в себя все составляющие целых мышц (например, жировую и соединительную ткани). У крыс, расхождение в волокнах и размерах мышцы увеличивается с перегрузкой. В связи с этим было высказано предположение, что мышечные волокна развиваются за счет экстрацеллюлярного компатрмента [138]. Необходимо также отметить, что в большинстве исследований на человеке с использованием техники биопсии мышц найдена большая гипертрофия, чем в тех, в которых используется измерение анатомического поперечника. [11,45,68,139].
Исследования плотности упаковки миофиламентов показали, что она очень постоянна до и после тренировки [52, 134]. Большинство современных исследований показало, что напряжение мышечных волокон, разделенных на типы в зависимости от изоформы тяжелых цепей миозина, не изменяется после 12 недель гипертрофической силовой тренировки. [72,140,141] и аналогично для лиц, выполняющих сидячую работу и тренировку с сопротивлением более 6 лет [142]. Таким образом, нет никаких доказательств адаптации плотности поперечных мостиков или внутренних сократительных свойств скелетных мышц (специфических напряжений) после гипертрофической силовой тренировки.
1.5.3. Сухожилия и соединительная ткань
Скелетные мышцы окутаны соединительной тканью, которая могут играть роль в передаче усилий сухожилиям [143] и рабочая гипертрофия, как известно, увеличивает синтез коллагена в мышцах животных [144]. Однако, есть данные, о фиксированной доли соединительной ткани в скелетных мышцах, несмотря на их гипертрофию (≈ 13% у бодибилдеров и лиц, не занимающихся спортом [124]), хотя это не исключает возможность некоторой пластичности соединительной ткани. Расположение соединительной ткани относительно отдельных мышечных волокон, может влиять на производство силы. Например, если прикрепление соединительной ткани осуществляется между сухожилием и промежуточной частью мышечных волокон, то площадь поперечного сечения волокна будет расти [145]. По сути, одно продольное волокно с дополнительным сухожильным прикреплением на половине его длины может, по сути, действовать с силой, эквивалентной двум параллельным волокнам. Действительно ли это происходит неизвестно, но теоретически, это можно было бы проверить, так как это может вызвать существенное воздействие на мышечную механику. Увеличение жесткости сухожилия у животных было обнаружено в ответ на нагрузку [146,147], а у людей – после изометрической [148] и изотонической гипертрофической тренировки [149, 150]. Reeves et al. [150] обнаружили 65% и 69% увеличение жесткости сухожилия коленной чашечки и модуля Юнга соответственно, после 14 недель тренировки для мышц-разгибателей голени. Жесткость сухожилия влияет на время, необходимое, чтобы растянуть последовательный упругий компонент и поэтому влияет как на электромеханическую задержку, так и на скорость нарастания силы [151]. От этого зависит быстрота применения силы. Увеличенная жесткость также уменьшает удлинение сухожилия и, скорее всего, изменяет характеристики «длина-сила» тренированных мышц, хотя этот вопрос не был специально исследован. Последние исследования поперечного сечения сухожилий показали большую толщину сухожилий у спортсменов, выполняющих силовые тренировки по сравнению с контрольной группой [152]. У животных, бегающих с высокой скоростью, была обнаружена гипертрофия сухожилия [153, 154]. Однако, длительные исследования на людях (14 недель гипертрофической силовой тренировки) этого не подтвердили [149, 150], возможно, потому, что это слишком короткий период. Однако двухфазный ответ с начальной атрофией и последующей гипертрофией наблюдается у сухожилий свиней в ответ на упражнения на выносливость. [147, 155]. Структурные изменения внутри сухожилия в ответ на гипертрофическую силовую тренировку у людей не исследовались, однако, исследования на животных показывают, что увеличенный диаметр и плотность упаковки коллагеновых волокон, а также изменения в структуре коллагена (волнистость фибриллы) [156, 157] могут повлиять на жесткость сухожилия. В то время как доля соединительной ткани в скелетных мышцах не меняется при гипертрофической силовой тренировке, изменения в расположении соединительных тканей неизвестны. Существуют убедительные доказательства, что увеличение жесткости сухожилия, вероятно, связано с рядом структурных изменений, и гипертрофия сухожилия также наступает после достаточно долгого периода тренировки.
1.5.4. Мышечная архитектура
Направление мышечных волокон относительно соединительной ткани и сухожилия – это релевантное соединение механики, которое влияет на силу мышц и может показать степень пластичности при HRST. Когда угол перистости мышечного волокна (AoP) увеличивается, возрастает упаковка мышечных волокон при одном и том же значении анатомического поперечника (существенно возрастает физиологический поперечник), но соединительной ткани передается меньше усилия из-за возрастания угла перистости. Поэтому, влияние AoP на силу – это разъединение этих двух факторов (упаковки против механических недостатков). Alexander и Vernon [152] сосчитали, что сила, производимая мышцей при закрепленных концах пропорциональна синусу удвоенного угла перистости. В соответствии с этой зависимостью, угол перистости в 45 градусов является оптимальным. В то время как большинство мышц имеют волокна, которые направлены вдоль общей линии действия, некоторые из них прикрепляются под углом к общей линии действия. Поэтому любое увеличение угла перистости увеличивает силу, даже, если не происходит увеличения анатомического поперечника.
В большом количестве работ обнаружена связь между разными показателями размера мышц и углом перистости на большом количестве тренированных и нетренированных испытуемых [159-161]. Можно предположить, что при гипертрофии увеличивается угол перистости мышечных волокон. В более ранних исследованиях [162] изменения в угле перистости латеральной широкой мышцы бедра (VL) после 12 недель тренировок не были найдены, хотя эти авторы допускали, что чувствительность их ультразвуковой измерительной техники могла быть недостаточной, чтобы зарегистрировать изменения угла перистости волокна. Aаgard et al. [11], было сообщено об увеличении угла перистости в латеральной широкой мышце бедра с 8 до 10,7 градусов (+36%) после 14 недель HRST. Увеличение угла перистости способствовало возрастанию физиологического поперечника мышцы (PCSA) и, таким образом, ощутимо могло увеличить изометрическую силу (+16%) чем анатомический поперечник (ACSA) или объем мышц (+10%). После 10-ти недель HRST трехглавой мышцы плеча было обнаружено увеличение угла перистости мышечных волокон на 17,0-19,2 градусов или + 16% [163] , а после и 16-ти недель HRST – на 16,5-21,3 градусов или на + 29% [164]. Reeves et al. [165] нашли, что после 14 недель HRST угол перистости латеральной широкой мышцы бедра увеличился от 28 до 35 %, в зависимости от угла в коленном суставе. Что более уникально, эти авторы измерили также угол перистости во время максимального изометрического сокращения, находя увеличение на 10-16% как результат тренировки.
Эти недавние исследования обеспечивают доказательство того, что угол перистости (AoP) увеличивается под воздействием гипертрофической силовой тренировки (HRST) и, так как большинство мышц имеют AoP, реально ниже оптимальных 45 градусов, этим предполагается основательный вклад в увеличение силы.
2. АДАПТАЦИЯ НЕРВНОЙ СИСТЕМЫ
Адаптация нервной системы к тренировкам с большими отягощениями (resistance training) – является очень важной из-за специфического характера адаптации при развитии силы, а также очевидного повышения напряжения мышцы после периода спортивной тренировки. В отличие от морфологической адаптации, существуют значительные противоречия в объяснении природы неврологических изменений, которые сопровождают тренировку на развитие силы. До недавнего времени большая часть доказательств неврологической адаптации имела только косвенные подтверждения, которая могла быть подвергнута сомнению методологически или нейрофизиологическим образом. Эти методологические неточности остаются до сих пор. Недавние исследования очертили круг определенных нервных механизмов, вносящих свой вклад в увеличение силы, вызванное тренировкой.
Sale et al. [166] сравнил проявление произвольной силы с навыком, в котором агонисты должны быть максимально активированы, при минимальной активации антагониста. Нервная адаптация представляет по существу изменения в координации и обучении, которые облегчают улучшенное рекрутирование и активацию мышц во время выполнения определенной задачи на развитие силы мышцей.
2.1. Непрямые доказательства нервной адаптации. Обучение и координация
Непропорционально большое увеличение силы мышц, по сравнению с их размерами, особенно на ранних стадиях силовой тренировки, является косвенным доказательством влияния нейрогенных факторов. Однако, как обсуждалось в первом разделе, многочисленные морфологические изменения могли также составлять это повышение силы (например, изменения в архитектуре мышечных волокон, параллельном и последовательном упругих компонентах, типе мышечных волокон). Пока некоторые исследователи, особенно Aagaard et al. [11] пытались включить вклад некоторых из этих факторов, чтобы вычислить изменения в мышечном волокне в естественных условиях после тренировки, Gandevia [167] указал, что очень трудно оценить совокупные эффекты этих необходимых исправлений.
2.1.1. Специфика тренировочной адаптации
Другие, хотя и косвенные, но более убедительные признаки реальной нервной адаптации могут быть взяты из наблюдений, сделанных во время многочисленных исследований силовых тренировок. В частности, увеличение динамической силы в преодолевающем режиме при выполнении упражнений с максимальными отягощениями (1 RM – 1 повторный максимум) значительно больше, чем увеличение изометрической силы [65, 168]. Несомненно, такие результаты значительно облегают изучение специфики тренировочной задачи. Определенная доля этой задачи связана со спецификой положения тела, которая зависит от поставленной задачи. Поскольку человеческое тело — взаимосвязанная механическая система, необходимо найти правильное положение конечностей и установить опору до начала силовой нагрузки [169]. Развитие силы и мощности после тренировки зависит от принимаемых положений [170], степени фиксации мышц и частоты их сокращения, данные показатели могут отличаться даже для аналогичных, по внешнему виду упражнений [168]. В своей последней работе Nozaki et al. [171] обращает внимание на изменчивость межмышечной координации и работы смежных суставов, как между, так и внутри субъектов, подвергающихся воздействию, даже во время, казалось бы, простых односуставных упражнений (например, разгибание голени в коленном суставе). Тем самым подтверждается тот факт, что простые на первый взгляд действия, очевидно, требуют определенных навыков применения силы.
2.1.2. Перекрестный тренировочный эффект
Есть все основания полагать, что имеет место перекрестный эффект при тренировке одной конечности, в результате чего происходит увеличение силы на противоположной, не тренируемой конечности [172-174] (обзор представлен Zhou [175]). Это подтверждает предположение о центральной адаптации при тренировочном воздействии на организм человека [176]. Однако, в некоторых исследованиях перекрестный эффект не обнаружен [3, 136, 177]. Можно предложить, что перекрестный тренировочный эффект возникает частично вследствие стабилизации или фиксации активности не тренируемой конечности во время упражнения [178], при этом электроактивность (EMG) противоположной мышцы составила всего 15 % от нагрузки, зарегистрированной во время максимального произвольного сокращения работающей мышцы (MVC) [179]. Очевидно, что вклад тренируемых мышц-синергистов, несмотря на попытки изолировать группу мышц во время снятия силовых замеров, может способствовать увеличению силы нетренируемой конечности.
Начальная стадия силовой тренировки должна формировать правильную модель межмышечной координации (то есть мышц-стабилизаторов, синергистов и антагонистов) [168], после чего, она может быть опробована, например, на противоположной стороне [167]. В подтверждение вышеизложенного имеются данные о том, что перекрестные тренировочные эффекты наблюдаются при работе мышц с определенной скоростью [180, 181]. Длительность такого предварительного обучения, вероятнее всего, зависит от первоначального уровня физической активности и координации (навыков) участников тренировочной задачи, и является вероятным объяснением разнообразных результатов при изучении перекрестных эффектов. Последние данные показали, что перекрестные эффекты могут проявляться вне зависимости от общей тренированности и координации, и включать изменения активации агониста. Используя технику вызванных сокращений (ITT) Shima et al. [182] зафиксировали существенное увеличение активации агонистов в тренируемой и контрлатеральной конечности спустя шесть недель тренировок.
2.1.3. Представление о сокращении мышц
В некоторых мышцах представление об их сокращении вызывают прирост силы, что означает адаптацию центральной нервной системы. Подобные эксперименты на сгибателе мизинца [183], внутренней стороне мышц руки, сгибателях спины [185] показали, увеличение силы, как от реальной нагрузки, так и при представлении о сокращении, которые были выше, чем у контрольной группы. Совсем недавно Zijdewind et al. [184] сравнивал результаты влияния семи недель представлений о сокращениях мышц, нагрузки низкой интенсивности и контрольной группы, которая не тренировалась на момент силы сгибателей стопы. Эти авторы обнаружили значительно больший прирост силы в группе, практикующей представления о сокращениях (36%) по сравнению с группой низкой интенсивности (13%) и контрольной группой (14%). Напротив, Herbert et al. [186] применили эту же идею к сгибателям плеча и обнаружили, что представления о сокращениях мышц дали тот же эффект, что и в контрольной группе (не тренирующихся) и были значительно ниже, чем в группе реально тренирующихся испытуемых. Это может иметь место по той причине, что возможности максимальной активации сгибателей плеча значительно выше, чем у других групп мышц [187], и, следовательно, они имеют значительно меньше возможностей для адаптации за счёт центральной нервной системы. Несмотря на то, что дальнейшие исследования, безусловно, необходимы, в целом это свидетельствует о наличии существенного прироста силы основных мышечных групп при тренировке с представлением о сокращениях без морфологических адаптационных изменений. Т.е. поддерживается роль центральных корковых адаптационных изменений в ответ на регулярные тренировки с отягощениями.
2.2. ИЗМЕНЕНИЯ В АКТИВАЦИИ МЫШЦ-АГОНИСТОВ?
Тот простой факт, что даже при максимальных сокращениях показатели силы могут сильно варьировать, нужен для того, чтобы показать, что истинной максимальной силы, очень трудно достичь [167]. Более того, бытует мнение о том, что здоровый, но нетренированный индивид не может в достаточной степени активировать мышцы при максимальном произвольном сокращении, даже при значительной мотивации [188,189]. При использовании упражнений с отягощениями активация мышц-агонистов может быть увеличена за счёт рекрутирования большего количества ДЕ, а также увеличения их частоты импульсации, при условии, что эти переменные были субмаксимальными до тренировок.
2.2.1. Электромиография
Данные поверхностной электромиографии (SEMG) использовались многими исследователями в попытке оценки изменений в активации мышц-агонистов. Огромное количество исследований свидетельствует о том, что поверхностная электромиограмма (SEMG) мышц-агонистов достоверно возрастает при силовой тренировке, особенно в первые 3-4 недели, что было принято как доказательство изменений в нервной активации мышц [33,46,48,172,173,190,191]. Hakkinen и Komi [190] обнаружили, что изменения в SEMG тесно связаны с изменениями в силе в течение 16 недель тренировки и 8 недель отдыха (рис. 5).
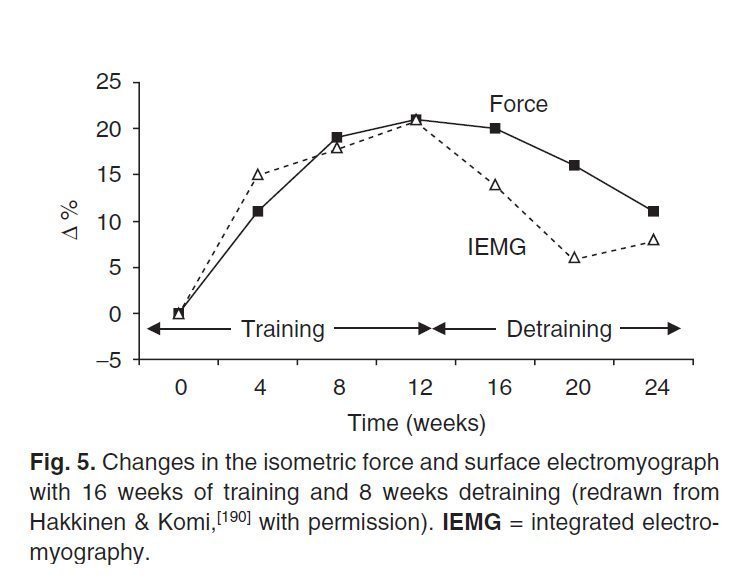
Рис.5. изменения в изометрической силе и поверхностной ЭМГ после 16 недель тренировки и 8 недель отдыха (по Hakkinen и Komi [190] с разрешения авторов) IEMG интегрированная электромиограмма
Напротив, некоторые исследования не выявили никаких изменений в поверхностной электромиограмме после тренировки [3, 8, 192, 193]. Чтобы проанализировать факторы, ответственные за резкий скачок в силе в начале программы тренировок, Holtermann et al. [194] наблюдал изменения в мышцах спины и электромиографию большеберцовой мышцы посредством больших сетчатых электродов, после девяти тренировочных занятий в течение пяти дней. В то время как сила увеличилась на 16%, амплитуда пиков SEMG понизилась на 11%. Расхождения в результатах SEMG можно объяснить рядом проблем, связанных с интерпретацией результатов этого исследования.
Технические проблемы с данными измерениями широко известны, и в то время как технологии электромиографии совершенствуются, все же вопрос о воспроизводимости результатов измерений остаётся открытым. Проблемы расположения электродов, различное суммарное сопротивление кожи и подкожной клетчатки, в купе с изменениями в морфологии мышц, как правило, весьма усложняют задачу вычленения долгосрочных изменений в данных SEMG.
Интерпретация увеличения SEMG как отражение повышения нервной активации также рассматривается как значительное упрощение. Во-первых, картина SEMG изменяется при изменениях в синапсах, передающих возбуждение мышцам, в частности, изменения потенциала действия мышечного волокна [167]. Большое количество факторов изменяется в ходе силовой тренировки, которые могут изменить потенциал действия одного волокна, в том числе: тип волокна, размер волокна, мембранный потенциал [195], внутримышечные концентрации ионов и содержание калий-натриевого насоса [196,197]. Во-вторых, крупные ДЕ расположены в большей степени у периферии мышц, ближе к коже [58,198], и любые изменения в их деятельности могут иметь преувеличенное влияние на данные SEMG. Суммарное воздействие этих факторов, а также изменчивость электрического импеданса, может быть проконтролировано/нормализовано измерением потенциала действия мышцы (М-волна, М-рефлекс), производимым сверхмаксимальным стимулированием нерва. Было обнаружено возрастание пиков электромиограммы, в то время как амплитуда М-рефлекса оставалась постоянной [199,200]. Другими исследователями было выявлено параллельное возрастание пиков электромиограммы и амплитуды М-рефлекса [201].
Наконец, в то время как увеличение SEMG может отражать увеличение количества задействованных волокон или частоту разрядов, суммарная ЭМГ также чувствительна к изменению в синхронизации разрядов ДЕ. Вне фазы суммарной SEMG может привести к отмене потенциалов действия двигательной единицы, что не обязательно отражает любые изменения в активации (возможные изменения в синхронизации рассматриваются в разделе 2.3.2).
2.2.2. Тетаническая стимуляция
Максимальная мышечная активация была вычислена различными способами, но в основном при условии относительной изоляции (например, односторонние изометрические упражнения, подразумевающие работу одной конечности). Сверхмаксимальные тетанические стимуляции, как представляется, являются наиболее полным методом оценки уровня произвольной активации мышц, хотя отсутствие активации синергистов и стабилизаторов ставит под сомнение обоснованность такого подхода. Из-за соответствующих трудностей, было проведено мало исследований. Было обнаружено, что изометрическая сила при максимальном произвольном сокращении (MVC) соответствовала силе, создаваемой тетанической стимуляцией у нетренированных субъектов [202 — 204], хотя точность данных ранних исследований сомнительна. После периода тренировки сравнение изменений в произвольной и вызванной электрической стимуляцией силе мышц, также было использовано для выяснения важности произвольной активации в развитии силы. Однако, доказательства сомнительны, учитывая данные о том, что в одних исследованиях показано, что тренировка с произвольными сокращениями способствует возрастанию [199,205] силы, а в других – она не оказывает никакого воздействия [206,207] на силу, полученную в результате электрической стимуляции. Третьим направлением исследований было сравнение тренировочного эффекта от электрической стимуляции и от произвольных сокращений. В ряде исследований использовался метод электрической стимуляции при тренировке, и в одних было заявлено, что сила достоверно взросла [208,209], в других – что она возросла в той же степени, как и от обычной тренировки [205, 210, 211], а по некоторым данным сила и анатомически поперечник мышцы возросли после стимуляции больше, чем после обычной тренировки [212]. Это свидетельствует о том, что значительный прирост в силе возможен и без вовлечения центральной нервной системы.
2.2.3. Интерполированная техника вызванных сокращений
Интерполированная техника вызванных сокращений (ITT) активно используется для измерения уровня мышечной активации [213-215]. В многочисленных исследованиях были использованы различные формы вызванных сокращений для того, чтобы заключить, что здоровые нетренированные люди могут достичь максимальной активации при изометрических усилиях [167]. Существует более широкое признание важности ряда технических и методологических вопросов в использовании этой техники (см. Folland и Williams [216] и Shield и Zhou [217]). Максимум нервной активации зависит от специфики мышцы [214], например, сгибатели плеча активируются лучше, чем четырёхглавая мышца бедра [187]. Примечательно, что более поздние работы свидетельствует о том, что активация многих групп мышц, редко бывает максимальной. Это доказывается тем, что активация четырехглавой мышцы бедра во время изометрических MVC составляет 85-95% у здоровых, нетренированных людей [182, 218-221]. В то время в большинстве более старых исследований не обнаружено увеличения произвольной активации после тренировки с отягощениями, [136, 177, 222], в более поздних исследованиях было обнаружено повышение активации после тренировки [165,182, 223, 224]. Еще одной разработкой в этой области является предположение, что максимум активации мышц во время изометрических усилий вполне может быть зависим от специфического угла. /h3br /B/h4ecker & Awiszus [225] нашли, что степень активации четырехглавой мышцы бедра при угле 40° в коленном суставе на 20% меньше, чем при 90 ° в коленном суставе (рис. 6а), и эти данные были недавно подтверждены [226].
2.2.4. Динамическая активность мышц
Большое число авторов предположили, что во время медленных концентрических сокращений, характерных для подходов на максимальное усилие, уменьшается нервная активация [189, 228, 229]. Используя электромиографию, Aagaard et al. [15] нашел подавление нервной активации во время максимальных медленlaquo;длина-силаных концентрических сокращений, которая была частично отменена после 14 недель занятий с отягощениями. Исследования с использованием наложения раздражающих электродов, как правило, опровергают эту теорию [230, 231]. Однако, используя ITT, Babault et al. [227] нашли, что активация будет значительно ниже для медленных концентрических, чем для изометрических сокращений (89,7% против 95,2%, соответственно) [рис. 6b].
Рис.6. Прямые доказательства, полученные посредством техники вызванных сокращений того, что способность к максимальной активации мышц-агонистов варьирует от угла в суставе /длины мышцы (а) данные Becker and Awiszis (225) и типа мышечного сокращения (b) данные Babault et al (227) с разрешения авторов.
Есть немало свидетельств субмаксимальной нервной стимуляции у неподготовленных испытуемых во время эксцентрических сокращений. Эксцентрическая часть зависимости «сила-скорость» для нетренированных людей, полученная в естественных условиях, пок 2.1.2. Перекрестный тренировочный эффектазывает заметное различие по сравнению с зависимостью, полученной в лабораторных условиях. В частности, сила не увеличивается при удлинении (эксцентрической активности) по сравнению с изометрической активностью [232]. Примечательно, что это несоответствие отсутствует для очень хорошо подготовленных лиц [232, 233] и исчезает при электрической стимуляции нетренированных людей [234]. Кроме того, эксцентрическая тренировка ранее нетренированных лиц приводит к значительно большему возрастанию в эксцентрически-специфической силе и электромиограф/h4ии, чем концентрическая тренировка концентрической силы [235]. Взятые вместе, эти данные достоверно указывают на сбой в активации мышц во время максимальных эксцентрических сокращений у нетренированных людей, либо из-за плохой супраспинальной активации или, что более вероятно, спинного ингибирования посредством афферентов (например, группы Ib-афферентов рецепторов Гольджи, афферентов группы Ia, группы II и III группы мышечных веретен и клеток Реншоу), хотя точный механизм остается неизвестным [15]. Существует всё больше доказательств того, что у нетренированных, но здоровых людей есть возможность для увеличения нервной активации синергистов. Величина этого центрального резерва, и, следовательно, потенциал для улучшения профессиональной подготовки, вероятно, будет зависеть от группы мышц, расp style=»text-align: justify;» 2.2. ИЗМЕНЕНИЯ В АКТИВАЦИИ МЫШЦ-АГОНИСТОВ? сматриваемого типа мышц, длины мышц и положения конечностей, а также сложности и знакомства с двигательной задачей (то есть двусторонней деятельности или совместной деятельности нескольких конечностей).
2.3. СПЕЦИФИЧЕСКИЕ МЕХАНИЗМЫ МЫШЕЧНОЙ АДАПТАЦИИ
Увеличение активации агонистов мышц после HRST могло произойти из-за возрастания рекрутирования ДЕ или частоты импульсации. Во время медленного сокращения после отдыха вклад этих двух факторов в увеличение активации очень зависит от рассматриваемой мышцы. У больших мышц, возможно больше проявляются процессы рекрутирования, чтобы достигнуть высокh3их уровней произвольной силы [236, 237]. Достоверное доказательство увеличения рекрутирования ДЕ под воздействием тренировки потребовало бы демонстрации ранее не вовлеченных в работу ДЕ, которые рекрутировались после тренировки. К сожалению, это вне возможности методов, которые применяются в настоящее время. Ясно, что оба способа: увеличение рекрутирования и/или возрастание частоты требует вовлечения некоторых форм нервной регуляции на спинном или супраспинальном уровне.
2.3.1. Частота импульсации
Используя большой сетчатый электрод, Holtermann et al. [194] оценили изменения в средней частоте SEMG (поверхностной ЭМГ) после 9 тренировочных занятий со сгибателями спины. Они не нашли изменений в средней частоте, которая расценивается как мера рекрутирования ДЕ, [238] несмотря на 16%-ое увеличение силы. Техника записи внутримышечных потенциалов ЭМГ, позволяет точно исследовать частоту импульсации ДЕ (MUFF) у людей в естественных условиях. MUFF может быть намного выше в течение очень кратких периодов (первые три импульса) в начале максимального произвольного усилия (100–200Hz [200]), однако частота импульсации может значительно снизиться в момент генерации максимального уровня силы (20–30Hz) [236,237,239,240]. Любопытно, что при непроизвольной стимуляции отношение «сила-частота», наблюдаемое у ДЕ мышц человека, предполагает, что уровень их импульсации должен быть, по крайней мере, в 50 Гц, чтобы достигнуть максимального значения силы при тетаническом сокращении [241,242]. Взятое в изоляции, это могло бы подтвердить значительную способность к увеличениям, возможно до 2-кратного, в MUFF во время максимальных произвольных сокращений, способствуя увеличенной силе после тренировки. Однако, считается, что явления, такие как подобные резкому повышению силы на единичный стимул в начале тетанического сокращения [243] и вызванные потенциалы [244] могут вызвать большее производство силы на более низких частотах, чем ожидаемые. Начальная короткая, высокочастотная импульсация в 2–4 импульса в начале сокращения увеличивает последующее производство силы и известна как catch-like способность скелетной мышцы [243]. Twitch потенциалы также относятся к большему сократительному ответу на единственный пульс после деятельности мышц, что может облегчить тетанические сокращения на более низких частотах иннервации. Во время максимальной генерации силы MUFF, как находили, была выше в мышцах у пауэрлифтеров пожилого возраста по сравнению с нетренируемыми лицамnbsp;несмотря на их гипертрофию ( Направление мышечных волокон относительно соединительной ткани и сухожилия /pи соответствующего возраста (23,8 Гц против 19,1 Гц, соответственно) [245]. Два длительных по времени исследования нашли увеличение MUFF после HRST [174, 200]. Van Cursem et al. [200] нашли у тренируемых в течение 12 недель (60 учебных сессий) посредством быстрых, баллистических сокращений, более раннюю активацию ДЕ, экстра дублеты (дуплеты) и увеличенную MUFF в начале баллистических сокращений после тренировки. Возможно эта адаптация будет способствовать увеличению в скорости развития силы и ускорения во время быстрых динамических сокращений, однако эффект их влияния на уровень MUFF и силы в момент максимального медленного произвольного сокращения неизвестен. Patten et al. [174] не сообщил ни о каком эффекте влияния на MUFF после двух недель силовой тренировки. В этом исследовании самые большие изменения (в силе и MUFF) произошли между двумя основными тестами, возможно из-за незнакомой природы движения (отведение 5-го пальца), небольшого числа испытуемых и короткой продолжительности обучения.
2.3.2. Синхронизация
Синхронизация измеряется уровнем корреляции между временами потенциалов действия активных ДЕ. ДЕ спортсменов, занимающихся силовыми упражнениями, как правило, более синхронизированные, чем у неподготовленных людей. Силовые тренировки, так же, по всей видимости, усиливают синхронизацию ДЕ [246, 247]. Однако не ясно, как повышение синхронизации может развивать силу [51, 176], т.к. при частоте импульсации, эквивалентной максимальному произвольному сокращению в развитии силы эффект синхронизации не проявляется [248,249].
2.3.3. Кортикальные адаптации
Методы нейровизуализации и транскраниальной магнитной стимуляции были протестированы на людях, тренирующихся в режиме низкой мышечной активности, чтобы вызвать изменения в первичной моторной коре, такие как: формирование двигательных представлений и увеличение корковой или кортикоспинальной возбудимости для специфичных мышц и движений. Подобное привыкание может также помочь объяснить, как воображаемые тренировки/ментальные практики могут повысить силу человека. Однако более детальные исследования с использованием методов транскраниальной стимуляции в ответ на повышение силы обнаружили неожиданный спад в кортикоспинальной возбудимости первой дорсальной межкостной и двуглавой мышцы плеча, что ставит под сомнение любую значительную корковую адаптацию.
2.3.4. Спинальные рефлексы
Афферентная обратная связь в форме спинальных рефлексов во время сокращения могла увеличить или уменьшить воздействие супраспинальных воздействий на мышцу. Были исследованы вызванные спинальные рефлексы, чтобы изучить любые изменения в мотонейронах спинного мозга после HRST, особенно их чувствительность к афферентной обратной связи. Рефлекс Хоффмана (или H-рефлекс) является искусственно вызванным рефлексом, который используется, чтобы проверить эффективность передачи стимула, поскольку это проходит от афферентных волокон через мотонейронный пул к двигательным волокнам. Как предполагается, это дает приблизительную оценку возбудимости мотонейронного пула [260]. V-волны — электрофизиологический вариант H-рефлекса, но будучи зарегистрированы во время максимального произвольного сокращения (MVC),они могут отразить моторную деятельность нейронов [261]. Ответ на H-рефлекс был измерен в покое, и он не изменился после тренировки [223], хотя уместность этого измерения была подвергнута сомнению [261]. Во время максимальных произвольных изометрических сокращений Sale et al. измерили V1 и V2 волны после тренировки, и сообщили, что нет никакого потенцирования [262] и существенного увеличения [166]. В недавнем исследовании Aagaard et al. [261] была тщательно оценена амплитуда M-волны (М-рефлекса), даже во время максимальных сокращений. Эти авторы нашли, что после 14 недель HRST наблюдалось 20%-ое увеличение изометрической силы, которое сопровождалось увеличенными амплитудами V-волны и H-рефлекса (55% и 19%, соответственно), рис.7.
Рис. 7. V-волна и амплитуда H-рефлекса (выраженные относительно максимального составного потенциала действия мышц [Mmax]) измеренные во время изометрических максимальных произвольных сокращений до и после 14 недель силовой тренировки (адаптированные данные Aagaard et al. [261] с разрешения авторов).
Увеличение амплитуды V-волны указывает на увеличенный поток от мотонейронов спинного мозга, который, как считают исследователи происходил наиболее вероятно из-за увеличенной частоты разрядов мотонойронов. Увеличение H-рефлекса после тренировки предполагает, что увеличение потока от мотонейронов было вызвано, частично, повышением их возбудимости, хотя большее увеличение V-волны по сравнению с H-рефлексом указывает на увеличенную супраспинальную активацию. Пока эти изменения убеждают, что они способствуют увеличению силы, хотя количественное функциональное значение этих эффектов остается неизвестным [263], и эти доказательства ясно противоречат удивительному уменьшению в кортикоспинальной возбудимости, которая наблюдалась после тренировки [258,259].
2.3.5. Коактивация антагонистов
Степень активации антагонистов во время любого упражнения зависит от широкого диапазона факторов, включая скорость и диапазон движения [264]. Любое сокращение антагонистов явно уменьшает уровень развития силы, но это также ослабляет, из-за реципрокной иннервации, возможность полностью активировать мышцы-агонисты. Исследования поперечного сечения нашли более низкую коактивацию в силе/мощности у тренируемых атлетов, чем у нетренируемых [265,266]. Carolan and Cafarelli [267] нашли существенное уменьшение в активации антагонистов, которая главным образом произошла на первой неделе изометрической программы тренировки четырехглавой мышцы бедра. После шести месяцев HRST четырехглавой мышцы бедра Hakkinen et al. [268] нашли уменьшение активации мышц задней поверхности бедра у стариков, но не нашли этого эффекта у лиц среднего возраста. Однако, другие исследования не нашли изменений в активации антагонистов после 9 недельных тренировок сгибателей спины (dorsiflexor) [194] или 14 недель тренировки разгибателей голени у пожилых людей [165]. Во время более сложных мультисоставных движений или всего тела уровень активации антагонистов может быть больше, возможно обеспечивая больше возможности для сокращения и тренировки.
3. ЗАКЛЮЧЕНИЕ
Увеличенной силе после HRST, как известно, способствует широкий диапазон морфологических и неврологических факторов. Увеличение размера мышц, выполняющих упражнения, как правило, расценивается как главная долгосрочная адаптация, хотя она очень вариативна между мышцами, подвергнутыми тренировке и вдоль их длины. Гипертрофия целой мышцы, кажется, линейно возрастает в течение первых шести месяцев тренировки и приписывается гипертрофии отдельных мышечных волокон посредством процессов гипертрофии и пролиферации миофибрилл, хотя гиперплазия может играть второстепенную роль. Может быть, увеличение отношения количества миоядер к объему цитоплазмы связано с супер регуляцией транскрипции или трансляции, но клетки-сателлиты активируются на очень ранних стадиях тренировки. Их быстрая пролиферация и соединение с существующими мышечными волокнами увеличивают число миоядер и это, по-видимому, является основой гипертрофии мышц. Гипертрофия мышечного волокна, как правило, больше в волокнах II типа и сопровождается увеличением угла волокна перистости, который способствует большему увеличению физиологического поперечника (PCSA) и производству силы, чем увеличение анатомического поперечника (ACSA). Вероятнее всего, что эти два фактора вносят основной вклад в увеличение силы всей мышцы и ее напряжения, несмотря на то, что для отдельного мышечного волокна напряжение не меняется. Влияние косвенных факторов (например, пересекающиеся эффекты, специфика задачи, быстрый прирост в силе в начале тренировки), и пока не категоричное влияние существенной нервной адаптации, которая может быть преобладающей при обучении и изменении в межмышечной координации агонистов, антагонистов и синергистов. Быстрое повышение силы в начале тренировки, в течение первых двух недель, которое происходит, прежде всего, из-за адаптации нервной системы, значительно увеличивает нагрузки и тренировочные стимулы, к которым мышца затем должна адаптироваться. Это помогает максимизировать дальнейшее увеличение силы, особенно морфологическую адаптацию, которая происходит, в то время тренировочного процесса. Более чувствительное использование интерполированной техники вызванных сокращений предполагает, что нетренированные люди могут быть не в состоянии полностью активировать мышцы-агонисты, и этот центральный резерв, кажется, зависит от диапазона специфических факторов задачи. Кроме того, пока спорное, исследование суммарной электрической активности (SEMG) указывает на увеличение активации агонистов после тренировки. Исследования, использующие трансчерепную стимуляцию, не нашли доказательств корковой или кортикоспинальной адаптации и противоречат исследованиям спинных рефлексов, которые указывают на увеличение супраспинального потока, и на возбудимость мотонейронов и вероятное увеличение MUFF после тренировки.
Подтверждения. В подготовке этого обзора не использовались никакие источники финансирования. У авторов нет конфликтов интересов, которые непосредственно относятся к содержанию этого обзора.
ЛИТЕРАТУРА
1. Morganti CM, Nelson ME, Fiatarone MA, et al. Strength improvements with 1 yr of progressive resistance training in older women. Med Sci Sports Exerc 1995; 27: 906-12
2. Paavolainen L, Paavolainen L, Hakkinen K, et al. Explosive strength training improves 5-km running time by improving running economy and muscle power. J Appl Physiol 1999; 86: 1527-33
3. Garfinkel S, Cafarelli E. Relative changes in maximal force, emg, and muscle cross-sectional area after isometric training. Med Sci Sports Exerc 1992; 24: 1220-7
4. Housh DJ, Housh TJ, Johnson GO, et al. Hypertrophic response to unilateral concentric isokinetic resistance training. J Appl Physiol 1992; 73: 65-70
5. Tracy B, Ivey F, Hurlbut D, et al. Muscle quality: II. Effects of strength training in 65- to 75-yr-old men and women. J Appl Physiol 1999; 86: 195-201
6. Abe T, De Hoyos D, Pollock M, et al. Time course for strength and muscle thickness changes following upper and lower body resistance training in men and women. Eur J Appl Physiol 2000; 81: 174-80
7. Engstrom CM, Loeb GE, Reid JG, et al. Morphometry of the human thigh muscles: a comparison between anatomical sec- tions and computer tomographic and magnetic-resonance images. J Anat 1991; 176: 139-56
8. Narici M, Hoppeler H, Kayser B, et al. Human quadriceps cross- sectional area, torque and neural activation during 6 months strength training. Acta Physiol Scand 1996; 157: 175-86
9. Alway SE, Grumbt WH, Stray-Gundersen J, et al. Effects of resistance training on elbow flexors of highly competitive bodybuilders. J Appl Physiol 1992; 72: 1512-21
10. Tracy B, Ivey F, Metter JE, et al. A more efficient magnetic resonance imaging-based strategy for measuring quadriceps muscle volume. Med Sci Sports Exerc 2003; 35: 425-33
11. Aagaard P, Andersen J, Dyhre-Poulsen P, et al. A mechanism for increased contractile strength of human pennate muscle in response to strength training: changes in muscle architecture. J Physiol 2001; 534: 613-23
12. Fukunaga T, Roy RR, Shellock FG, et al. Specific tension of human plantar flexors and dorsiflexors. J Appl Physiol 1996; 80: 158-65
13. Roman WJ, Fleckenstein J, Straygundersen J, et al. Adaptations in the elbow flexors of elderly males after heavy-resistance training. J Appl Physiol 1993; 74: 750-4
14. Keen DA, Yue GH, Enoka RM. Training-related enhancement in the control of motor output in elderly humans. J Appl Physiol 1994; 77: 2648-58
15. Aagaard P, Simonsen E, Andersen J, et al. Neural inhibition during maximal eccentric and concentric quadriceps contraction: effects of resistance training. J Appl Physiol 2000; 89: 2249-57
16. Bamman MM, Newcomer BR, Larson-Meyer DE, et al. Evalua- tion of the strength-size relationship in vivo using various muscle size indices. Med Sci Sports Exerc 2000; 32: 1307-13
17. Fukunaga T, Miyatani M, Tachi M, et al. Muscle volume is a major determinant of joint torque in humans. Acta Physiol Scand 2001; 172: 249-55
18. Maganaris CN, Baltzopoulos V, Sargeant AJ. Changes in Achilles tendon moment arm from rest to maximum isometric plantar flexion: in vivo observations in man. J Physiol 1998; 510: 977-85
19. Maganaris CN, Baltzopoulos V. Predictability of in vivo changes in pennation angle of human tibialis anterior muscle from rest to maximum isometric dorsiflexion. Eur J Appl Physiol Occup Physiol 1999; 79: 294-7
20. Narici MV, Binzoni T, Hiltbrand E, et al. In vivo human gastrocnemius architecture with changing joint angle at rest and during graded isometric contraction. J Physiol 1996; 496: 287-97
21. Wilmore JH. Alterations in strength, body composition and anthropometric measurements consequent to a 10-week weight training-program. Med Sci Sports Exerc 1974; 6: 133-8
22. Cureton KJ, Collins MA, Hill DW, et al. Muscle hypertrophy in men and women. Med Sci Sports Exerc 1988; 20: 338-44
23. Welle S, Totterman S, Thornton C. Effect of age on muscle hypertrophy induced by resistance training. J Gerontol A Biol Sci Med Sci 1996; 51: M270-5
24. Kadi F, Bonnerud P, Eriksson A, et al. The expression of androgen receptors in human neck and limb muscles: effects of training and self-administration of androgenic-anabolic steroids. Histochem Cell Biol 2000; 113: 9
25. Edwards RH, Young A, Hosking GP, et al. Human skeletal muscle function: description of tests and normal values. Clin Sci Mol Med 1977; 52: 283-90
26. Schantz P, Randall-Fox E, Hutchison W, et al. Muscle fibre type distribution, muscle cross-sectional area and maximal voluntary strength in humans. Acta Physiol Scand 1983; 117: 219-26
27. Neder JA, Nery LE, Silva AC, et al. Maximal aerobic power and leg muscle mass and strength related to age in non-athlethic males and females. Eur J Appl Physiol Occup Physiol 1999; 79: 522-30
28. Toft I, Lindal S, Bønaa KH, et al. Quantitative measurement of muscle fiber composition in a normal population. Muscle Nerve 2003; 28: 101-8
29. Asmussen E. Development patterns in physical performance capacity. In: Larsson L, editor. Fitness, health and work capacity: international standards for assessment. New York: Mac- Millan, 1974: 8
30. Hettinger T. Physiology of strength. Springfield (IL): CC Thomas, 1961
31. Brown CH, Wilmore JH. The effects of maximal resistance training on the strength and body composition of women athletes. Med Sci Sports 1974; 6: 174-7
32. Wells CL. Women, sport and performance: a physiological perspective. Champaign (IL): Human Kinetics, 1985
33. Hakkinen K, Kallinen M, Linnamo V, et al. Neuromuscular adaptations during bilateral versus unilateral strength training in middle-aged and elderly men and women. Acta Physiol Scand 1996; 158: 77-88
34. Roth SM, Ivey FM, Martel GF, et al. Muscle size responses to strength training in young and older men and women. J Am Geriatr Soc 2001; 49: 1428-33
35. Colliander EB, Tesch PA. Responses to eccentric and concentric resistance training in females and males. Acta Physiol Scand 1991; 141: 149-56
36. Lexell J, Downham DY, Larsson Y, et al. Heavy-resistance training in older scandinavian men and women: short- and long-term effects on arm and leg muscles. Scand J Med Sci Sports 1995; 5: 329-41
37. Weiss LW, Clark FC, Howard DG. Effects of heavy-resistance triceps surae muscle training on strength and muscularity of men and women. Phys Ther 1988; 68: 208-13
38. O’Hagan FT, Sale DG, MacDougall JD, et al. Response to resistance training in young women and men. Int J Sports Med 1995; 16: 314-21
39. Hubal MJ, Gordish-Dressman H, Thompson PD, et al. Variability in muscle size and strength gain after unilateral resistance training. Med Sci Sports Exerc 2005; 37: 964-72
40. Knapik JJ, Wright JE, Kowal DM, et al. The influence of US army basic initial entry training on the muscular strength of men and women. Aviat Space Environ Med 1980; 51: 1086-90
41. Delmonico MJ, Kostek MC, Doldo NA, et al. Effects of moderate velocity strength training on peak muscle power and movement velocity: do women respond differently than men? J Appl Physiol 2005; 99: 1712-8
42. Fiatarone MA, Marks EC, Ryan ND, et al. High-intensity strength training in nonagenarians: effects on skeletal-muscle. J Am Med Assoc 1990; 263: 3029-34
43. Harridge SDR, Kryger A, Stensgaard A. Knee extensor strength, activation, and size in very elderly people following strength training. Muscle Nerve 1999; 22: 831-9
44. Ivey F, Tracy B, Lemmer J, et al. Effects of strength training and detraining on muscle quality: age and gender comparisons. J Gerontol A Biol Sci Med Sci 2000; 55: B152-7
45. Hakkinen K, Newton RU, Gordon SE, et al. Changes in muscle morphology, electromyographic activity, and force production characteristics during progressive strength training in young and older men. J Gerontol A Biol Sci Med Sci 1998; 53: B415-23
46. Narici MV, Roi GS, Landoni L, et al. Changes in force, cross- sectional area and neural activation during strength training and detraining of the human quadriceps. Eur J Appl Physiol Occup Physiol 1989; 59: 310-9
47. Hakkinen K, Pakarinen A, Kraemer WJ, et al. Selective muscle hypertrophy, changes in EMG and force, and serum hormones during strength training in older women. J Appl Physiol 2001; 91: 569-80
48. Hakkinen K, Alen M, Kraemer WJ, et al. Neuromuscular adaptations during concurrent strength and endurance training versus strength training. Eur J Appl Physiol 2003; 89: 42-52
49. Kanehisa H, Funato K, Kuno S, et al. Growth trend of the quadriceps femoris muscle in junior olympic weight lifters: an 18-month follow-up survey. Eur J Appl Physiol 2003; 89: 238-42
50. McDonagh MJN, Davies CTM. Adaptive response of mammalian skeletal-muscle to exercise with high loads. Eur J Appl Physiol Occup Physiol 1984; 52: 139-55
51. Jones DA, Rutherford OM, Parker DF. Physiological changes in skeletal muscle as a result of strength training. Q J Exp Physiol Cogn Med Sci 1989; 74: 233-56
52. MacDougall JD, Elder GCB, Sale DG, et al. Effects of strength training and immobilization on human-muscle fibers. Eur J Appl Physiol Occup Physiol 1980; 43: 25-34
53. Viitasalo JT, Saukkonen S, Komi PV. Reproducibility of mea- surements of selected neuromuscular performance variables in man. Electromyogr Clin Neurophysiol 1980; 20: 487-501
54. Schantz P, Fox ER, Norgren P, et al. The relationship between the mean muscle fibre area and the muscle cross-sectional area of the thigh in subjects with large differences in thigh girth. Acta Physiol Scand 1981; 113: 537-9
55. Halkjaer-Kristensen J, Ingemann-Hansen T. Variations in single fibre areas and fibre composition in needle biopsies from the human quadriceps muscle. Scand J Clin Lab Invest 1981; 41: i391- 5
56. Blomstrand E, Celsing F, Friden J, et al. How to calculate human muscle fibre areas in biopsy samples: methodological considerations. Acta Physiol Scand 1984; 122: 545-51
57. Mahon M, Toman A, Willan PL, et al. Variability of histochemical and morphometric data from needle biopsy specimens of human quadriceps femoris muscle. J Neurol Sci 1984; 63: 85-100
58. Lexell J, Taylor CC. Variability in muscle fibre areas in whole human quadriceps muscle: how much and why? Acta Physiol Scand 1989; 136: 561-8
59. Gollnick PD, Matoba H. The muscle fiber composition of skeletal muscle as a predictor of athletic success: an overview. Am J Sports Med 1984; 12: 212-7
60. Campos GER, Luecke TJ, Wendeln HK, et al. Muscular adaptations in response to three different resistance-training regimens: specificity of repetition maximum training zones. Eur J Appl Physiol 2002; 88: 50-60
61. Staron RS, Malicky ES, Leonardi MJ, et al. Muscle hypertrophy and fast fiber type conversions in heavy resistance-trained women. Eur J Appl Physiol Occup Physiol 1990; 60: 71-9
62. Tesch PA. Skeletal-muscle adaptations consequent to long-term heavy resistance exercise. Med Sci Sports Exerc 1988; 20: S132-4
63. Thorstensson A, Hulten B, Dobeln WV, et al. Effect of strength training on enzyme-activities and fiber characteristics in human skeletal-muscle. Acta Physiol Scand 1976; 96: 392-8
64. Hakkinen K, Komi P, Tesch P. Effect of combined concentric and eccentric strength training and detraining on force-time, muscle fiber and metabolic characteristics of leg extensor muscles. Scand J Sports Sci 1981; 3: 50-8
65. Dons B, Bollerup K, Bondepetersen F, et al. Effect of weight- lifting exercise related to muscle-fiber composition and muscle cross-sectional area in humans. Eur J Appl Physiol Occup Physiol 1979; 40: 95-106
66. Houston ME, Froese EA, Valeriote SP, et al. Muscle performance, morphology and metabolic capacity during strength training and detraining: a one leg model. Eur J Appl Physiol Occup Physiol 1983; 51: 25-35
67. Goldspink G, Ward PS. Changes in rodent muscle-fiber types during postnatal-growth, undernutrition and exercise. J Physiol 1979; 296: 453-69
68. Frontera WR, Meredith CN, Oreilly KP, et al. Strength conditioning in older men: skeletal-muscle hypertrophy and improved function. J Appl Physiol 1988; 64: 1038-44
69. Fitts RH, McDonald KS, Schluter JM. The determinants of skeletal-muscle force and power: their adaptability with changes in activity pattern. J Biomech 1991; 24: 111-22
70. Stienen GJM, Kiers JL, Bottinelli R, et al. Myofibrillar ATPase activity in skinned human skeletal muscle fibres: fibre type and temperature dependence. J Physiol 1996; 493: 299-307
71. Bottinelli R, Pellegrino MA, Canepari M, et al. Specific contributions of various muscle fibre types to human muscle performance: an in vitro study. J Electromyogr Kinesiol 1999; 9: 87-95
72. Widrick JJ, Stelzer JE, Shoepe TC, et al. Functional properties of human muscle fibers after short-term resistance exercise training. Am J Physiol Regul Integr Comp Physiol 2002; 283: R408-16
73. Nygaard E, Houston M, Suzuki Y, et al. Morphology of the brachial biceps muscle and elbow flexion in man. Acta Physiol Scand 1983; 117: 287-92
74. Maughan RJ, Nimmo MA. The influence of variations in muscle-fiber composition on muscle strength and cross-sectional area in untrained males. J Physiol 1984; 351: 299-311
75. Grindrod S, Round JM, Rutherford OM. Type-2 fiber composition and force per cross-sectional area in the human quadriceps. J Physiol 1987; 390: P154
76. Aagaard P, Andersen JL. Correlation between contractile strength and myosin heavy chain isoform composition in human skeletal muscle. Med Sci Sports Exerc 1998; 30: 1217-22
77. Gur H, Gransberg L, vanDyke D, et al. Relationship between in vivo muscle force at different speeds of isokinetic movements and myosin isoform expression in men and women. Eur J Appl Physiol 2003; 88: 487-96
78. MacDougall JD, Sale DG, Moroz JR, et al. Mitochondrial volume density in human skeletal-muscle following heavy renbsp;leg muscle mass and strength related to age in non-athlethicsistance training. Med Sci Sports Exerc 1979; 11: 164-6
79. Morkin E. Postnatal muscle fiber assembly: localization of newly synthesized myofibrillar proteins. Science 1970; 167: 1499-501
80. Goldspink G. The proliferation of myofibrils during muscle fibre growth. J Cell Sci 1970; 6: 593-603
82. Goldspink G, Howells KF. Work-induced hypertrophy in exercised normal muscles of different ages and the reversibility of hypertrophy after cessation of exercise. J. Physiol. 1974; 239: 179-93
83. Goldspink G. Cellular and molecunbsp; lar aspects of adaptation in skeletal muscle. In: Komi PV, editor. Strength and power in sport. London: Blackwell Science, 1992: 211-229
84. Patterson S, Goldspink G. Mechanism of myofibril growth and proliferation in fish muscle. J Cell Sci 1976; 22: 607-16
85. Ashmore CR, Summers PJ. Stretch-induced growth in chicken wing muscles: myofibrillar proliferation. Am J Physiol 1981; 241: C93-7
86. Moss FP. The relationship between the dimensions of the fibres and the number of nuclei during restricted growth, degrowth and compensatory growth of skeletal muscle. J Anat 1968; 122: 555-63
87. Moss FP, Leblond CP. Satellite cells as the source of nuclei in muscles of growing rats. Anat Rec 1971; 170: 421-36
88. Burleigh IG. Observations on the number of nuclei within the fibres of some red and white muscles. J Cell Sci 1977; 23: 269-84
89. Eisenberg BR, Kennedy JM, Wenderoth MP, et al. Anonymous enlarge cellular and molecular biology of muscle development. New York: AR Liss, 1989: 460
90. Landing BH, Dixon LG, Wells TR. Studies on isolated human skeletal muscle fibers, including a proposed pattern of nuclear distribution anstrong/pd a concept of nuclear territories. Hum Pathol 1974; 5: 441-61
91. Schmalbruch H. Muscle regeneration: fetal myogenesis in a new setting. Bib Anat 1986; 29: 126-53
92. Allen RE, Merkel RA, Young RB. Cellular aspects of muscle growth: myogenic cell proliferation. J Anim Sci 1979; 49: 115-27
93. Bourke DL, Wylie SR, Theon A, et al. Myosin heavy chain expression following transfer into regenerating chicken muscle. Basic Appl Myol 1995; 5: 43-56
94. Allen DL, Monke SR, Talmadge RJ, et al. Plasticity of myonuclear number in hypertrophied and atrophied mammalian skeletal muscle fibers. J Appl Physiol 1995; 78: 1969-76
9nbsp;Physiol 1992; 73: 65-70p5. Rosenblatt JD, Parry DJ. Gamma irradiation prevents compensatory hypertrophy of overloaded mouse extensor digitorum longus muscle. J Appl Physiol 1992; 73: 2538-43
96. Rosenblatt JD, Parry DJ. Adaptation of rat extensor digitorum longus muscle to gamma irradiation and overload. Pflugers Arch 1993; 423: 255-64
97. Rosenblatt JD, Yong D, Parry DJ. Satellite cell activity is required for hypertrophy of overloaded adult rat muscle. Muscle Nerve 1994; 13. Roman WJ, Fleckenstein J, Straygundersen J, et al. Adaptations/p/p 17: 608-13
98. Kadi F, Eriksson A, Holmner S, et al. Cellular adaptation of the trapezius muscle in strength-trained athletes. Histochem Cell Biol 1999; 111: 189-95
99. Kadi F, Eriksson A, Holmner S, et al. Effects of anabolic steroids on the muscle cells of strength-trained athletes. Med Sci Sports Exerc 1999; 31: 1528-34
100. Kadi F, Thornell LE. Concomitant increases in myonuclear and satellite cell content in female trapezius muscle following strength training. Histochem Cell Biol 2000; 113: 99-103
101. Roth S, Martel G, Ivey F, et al. Skeletal muscle satellite cell characteristics in young and older men and women after heavy resistance strength training. J Gerontol. A p /pBio Sci 2001; 56: B240-7
102. Kadi F, Schjerling P, Andersen LL, et al. The effects of heavy resistance training and detraining on satellite cells in human skeletal muscles. J. Physiol 2004; 558: 1005-12
103. Crameri RM, Langberg H, Magnusson P, et al. Changes in satellite cells in human skeletal muscle after a single bout of high intensity exercise. J Physiol 2004; 558: 333-40
104. Hikida R, Staron R, Hagerman F, et al. Effects of high-intensity resistance training 24. Kadi F, Bonnerud P, Eriksson A, et al. The expression of 30. Hettinger T. Physiology of strength. Springfield (IL): CC on untrained older men: II. Muscle fiber characteristics and nucleo-cytoplasmic relationships. J Gerontol A Biol Sci Med Sci 2000; 55: B347-54
105. Sartorelli V, Fulco M. Molecular and cellular determinants of skeletal muscle atrophy and hypertrophy. Sci STKE 2004; 2004: re11
nbsp; /p 106. Reitsma W. Skeletal muscle hypertrophy after heavy exercise in rats with surgically reduced muscle function. Am J Phys Med 1969; 48: 237-58
107. Appell HJ. Muscular atrophy following immobilization: a re- view. Sports Med 1990; 10: 42-58
108. Edgerton VR. Morphology and histochemistry of the soleus muscle from normal and exercised rats. Am J Anat 1970; 127: 81-8
109. Carrow RE, Heusener WW, Van Huss D, et al. Exercise and the incidence of muscle fibre splitting. Br Assoc Sports Med 1973; 7: 39-41
110. Ho KW, Roy RR, Tweedle CD, et al. Skeletal muscle fiber splitting with weight-lifting exercise in rats. Am J Anat 1980; 157: 433-40
111. Gollnick PD, Timson BF, Moore RL, et al. Muscular enlargement and number of fibers in skeletal muscles of rats. J Appl Physiol 1981; 50: 936-43
112. Gonyea WJ, Sale DG, Gonyea FB, et al. Exercise induced increases in muscle fiber number. Eur J Appl Physiol 1986; 55: 137-41
113. Kelley G. Mechanical overload and skeletal muscle fiber hyperplasia: a meta-analysis. J Appl Physiol 1996; 81: 1584-8
114. Antonio J, Gonyea WJ. Skeletal muscle fiber hyperplasia. Med Sci Sports Exerc 1993; 25: 1333-45
115. Sjostrom M, Lexell J, Eriksson A, et al. Evidence of fibre hyperplasia in human skeletal muscles from healthy young men? A left-right comparison of the fibre number in whole anterior tibialis muscles. Eur J Appl Physiol 1991; 62: 301-4
116. Mauro M. Muscle regeneration. New York: Raven Press, 1979
117. Appell HJ, Forsberg S, Hollmann W. Satellite cell activation in human skeletal muscle after training: evidence for muscle fiber neoformation. Int J Sports Med 1988; 9: 297-9
118. Tesch PA, Larsson L. Muscle hypertrophy in bodybuilders. Eur J Appl Physiol 1982; 49: 301-6
119. Larsson L, Tesch PA. Motor unit fibre density in extremely hypertrophied skeletal muscles in man: electrophysiological signs of muscle fibre hyperplasia. Eur J Appl Physiol 1986; 55: 130-6
120. MacDougall JD, Sale DG, Elder GC, et al. Muscle ultrastruсtural characteristics of elite powerlifters and bodybuilders. Eur J Appl Physiol 1982; 48: 117-26
122. Alway SE, Grumbt WH, Gonyea WJ, et al. Contrasts in muscle and myofibers of elite male and female bodybuilders. J Appl Physiol 1989; 67: 24-31
123. McCall GE, Byrnes WC, Dickinson A, et al. Muscle fiber hypertrophy, hyperplasia, and capillary density in college men afppter resistance training. J Appl Physiol 1996; 81: 2004-12
124. MacDougall JD, Sale DG, Alway SE, et al. Muscle fiber number in biceps brachii in bodybuilders and control subjects. J Physiol 1984; 57: 1399-403
125. Burke RE, Kanda K, Mayer RF. The effect of chronic immobilisation on defined types of motor units in cat medial gastrocnemius. Science 1975; 174: 709-12
126. Walsh JV, Burke RE, Rymer WZ, et al. Effect of compensatory hypertrophy studied in individual motor units in medial gastrocnemius muscle of the cat. J Neurophysiol 1978; 41: 496-508
127. Haddad F, Qin AX, Zeng M, et al. Effects of isometric training on skeletal myosin heavy chain expression. J Appl Physiol 1998; 84: 2036-41
128. Hather BM, Tesch PA, Buchanan P, et al. Influence of eccentric actions on skeletal muscle adaptations to resistance training. Acta Physiol Scand 1991; 143: 177-85
129. Carroll TJ, Abernethy PJ, Logan PA, et al. Resistance training frequency: strength and myosin heavy chain responses to two and three bouts per week. Eur J Appl Physiol 1998; 78: 270-5
130. Schiaffino S, Gorza L, Sartore S, et al. Three myosin heavy chain isoforms in type 2 skeletal muscle fibres. J Muscle Res Cell Motil 1989; 10: 197-205
131. Andersen J, Aagaard P. Myosin heavy chain IIx overshoot in human skeletal muscle. Muscle Nerve 2000; 23: 1095-104
132. Williamson DL, Gallagher PM, Carroll CC, et al. Reduction in hybrid single muscle fiber proportions with resistance training in humans. J Appl Physiol 2001; 91: 1955-61
133. Andersen LL, Andersen JL, Magnusson SP, et al. Changes in the human muscle force-velocity relationship in response to resistance training and subsequent detraining. J Appl Physiol 2005; 99: 87-94
134. Claassen H, Gerber C, Hoppeler H, et al. Muscle filament spacing and short-term heavy-resistance exercise in humans. J Physiol 1989; 409: 491-5
135. Horber FF, Scheidegger JR, Gru¨nig BE, et al. Thigh muscle mass and function in patients treated with glucocorticoids. Eur J Clin Invest 1985; 15: 302-7
136. Jones DA, Rutherford OM. Human muscle strength training: the effects of three different regimens and the nature of the resultant changes. J Physiol 1987; 391: 1-11
137. Sipila¨ S, Suominen H. Effects of strength and endurance train- ing on thigh and leg muscle mass and composition in elderly women. J Appl Physiol 1995; 78: 334-40
138. Goldspink G. The combined effects of exercise and reduced food intake on skeletal muscle fibers. J Cell Comp Physiol 1964; 63: 209-19
139. Esmarck B, Andersen JL, Olsen S, et al. Timing of postexercise protein intake is important for muscle hypertrophy with resis- tance training in elderly humans. J Physiol 2001; 535: 301-11
140. Trappe S, Williamson D, Godard M, et al. Effect of resistance training on single muscle fiber contractile function in older men. J Appl Physiol 2000; 89: 143-52
141. Godard MP, Gallagher PM, Raue U, et al. Alterations in single muscle fiber calcium sensitivity with resistance training in older women. Pflugers Arch 2002; 444: 419-25
142. Shoepe TC, Stelzer JE, Garner DP, et al. Functional adaptability of muscle fibers to long-term resistance exercise. Med Sci Sports Exerc 2003; 35: 944-51
143. Street SF. Lateral transmission of tension in frog myofibers: a myofibrillar network and transverse cytoskeletal connections are possible transmitters. J Cell Physiol 1983; 114: 346-64
144. Goldberg AL, Etlinger JD, Goldspink DF, et al. Mechanism of work-induced hypertrophy of skeletal muscle. Med Sci Sports Exerc 1975; 7: 248-61
145. Huxley AF. Muscle structure and theories of contraction. Prog Biophysics Biophysical Chem 1957; 7: 255-318
146. Viidik A. Tensile strength properties of Achilles tendon systems in trained and untrained rabbits. Acta Orthop Scand 1969; 40: 261-72
147. Woo SL, Gomez MA, Amiel D, et al. The effects of exercise on coordinathe biomechanical and biochemical properties of swine digital flexor tendons. Biomech Eng 1981; 103: 51-6
148. Kubo K, Kanehisa H, Fukunaga T. Effects of different duration isometric contractions on tendon elasticity in human quadriceps muscles. J Physiol 2001; 536: 649-55
149. Kubo K, Kanehisa H, Fukunaga T. Effects of resistance and stretching training programmes on the viscoelastic properties of human tendon structures in vivo. J Physiol 2002; 538: 219-26
150. Reeves ND, Maganaris CN, Narici MV. Effect of strength training on human patella tendon mechanical properties of older individuals. J Physiol 2003; 548: 971-81
151. Bojsen-Møller J, Magnusson SP, Rasmussen LR, et al. Muscle performance during maximal isometric and dynamic contractions is influenced by the stiffness of the tendinous structures. J Appl Physiol 2005; 99: 986-94
152. Kongsgaard M, Aagaard P, Kjaer M, et al. Structural Achilles tendon properties in athletes subjected to different exercise modes and in Achilles tendon rupture patients. J Appl Physiol 2005; 99: 1965-71
153. Sommer HM. The biomechanical and metabolic effects of a running regime on the Achilles tendon in the rat. Int Orthop 1987; 11: 71-5
154. Birch HL, McLaughlin L, Smith RK, et al. Treadmill exercise- induced tendon hypertrophy: assessment of tendons with different mechanical functions. Equine Vet J Suppl 1999; 30: 222-6
155. Woo SL, Ritter MA, Amiel D, et al. The biomechanical and biochemical properties of swine tendons: long term effects of exercise on the digital extensors. Connect Tissue Res 1980; 7: 177-83
156. Wood TO, Cooke PH, Goodship AE. The effect of exercise and anabolic steroids on the mechanical properties and crimp morphology of the rat tendon. Am J Sports Med 1988; 16: 153-8
157. Michna H, Hartmann G. Adaptation of tendon collagen to exercise. Int Orthop 1989; 13: 161-5
158. Alexander RM, Vernon A. The dimensions of the knee and ankle muscles and the forces they exert. J Hum Movt Stud 1975; 1: 115-23
159. Kawakami Y, Abe T, Fukunaga T. Muscle-fiber pennation angles are greater in hypertrophied than in normal muscles. J Appl Physiol 1993; 74: 2740-4
160. Ichinose Y, Kanehisa H, Ito M, et al. Relationship between muscle fiber pennation and force generation capability in olympic athletes. Int J Sports Med 1998; 19: 541-6
161. Abe T, Brechue WF, Fujita S, et al. Gender differences in FFM accumulation and architectural characteristics of muscle. Med Sci Sports Exerc 1998; 30: 1066-70
162. Rutherford OM, Jones DA. Measurement of fibre pennation using ultrasound in the human quadriceps in vivo. Eur J Appl Physiol 1992; 65: 433-7
163. Kanehisa H, Nagareda H, Kawakami Y, et al. Effects of equivolume isometric training programs comprising medium or high resistance on muscle size and strength. Eur J Appl Physiol 2002; 87: 112-9
164. Kawakami Y, Abe T, Kuno SY, et al. Training-induced changes in muscle architecture and specific tension. Eur J Appl Physiol Occup Physiol 1995; 72: 37-43
165. Reeves ND, Narici MV, Maganaris CN. Effect of resistance training on skeletal muscle-specific force in elderly humans. J Appl Physiol 2004; 96: 885-92
166. Sale DG, MacDougall JD, Upton AR, et al. Effect of strength training upon motoneuron excitability in man. Med Sci Sports Exerc 1983; 15: 57-62
167. Gandevia SC. Spinal and supraspinal factors in human muscle fatigue. Physiol Rev 2001; 81: 1725-89
168. Rutherford OM, Jones DA. The role of learning and coordination in strength training. Eur J Appl Physiol 1986; 55: 100-5
169. Horak FB, Macpherson JM. Postural orientation and equilibrium. In: Rowell LB, Shepherd JT, editors. Handbook of physiology: section 12 exercise: regulation and integration of multiple systems. New York: Oxford University Press, 1996: 292
170. Wilson GJ, Murphy AJ, Walshe A. The specificity of strength training: the effect of posture. Eur J Appl Physiol Occup Physiol 1996; 73: 346-52
171. Nozaki D, Nakazawa K, Akai M. Uncertainty of knee joint muscle activity during knee joint torque exertion: the significance of controlling adjacent joint torque. J Appl Physiol 2005; 99: 1093-103
172. Komi PV, Viitasalo JT, Rauramaa R, et al. Effect of isometric strength training of mechanical, electrical, and metabolic aspects of muscle function. Eur J Appl Physiol Occup Physiol 1978; 40: 45-55
173. Moritani T, deVries HA. Neural factors versus hypertrophy in the time course of muscle strength gain. Am J Phys Med 1979; 58: 115-30
174. Patten C, Kamen G, Rowland D. Adaptations in maximal motor unit discharge rate to strength training in young and older adults. Muscle Nerve 2001; 24: 542-50
175. Zhou S. Chronic neural adaptations to unilateral exercise: mechanisms of cross education. Exerc Sport Sci Rev 2000; 28: 177-84
176. Sale DG. Neural adaptation to resistance training. Med Sci Sports Exerc 1988; 20: S135-45
177. Davies J, Parker DF, Rutherford OM, et al. Changes in strength and cross sectional area of the elbow flexors as a result of isometric strength training. Eur J Appl Physiol 1988; 57: 667-70
178. Young A, Stokes M, Round JM, et al. The effect of high- resistance training on the strength and cross-sectional area of the human quadriceps. Eur J Clin Invest 1983; 13: 411-7
179. Hortobagyi T, Scott K, Lambert J, et al. Cross-education of muscle strength is greater with stimulated than voluntary con- tractions. Motor Control 1999; 3: 205-19
180. Farthing JP, Chilibeck PD. The effect of eccentric training at different velocities on cross-education. Eur J Appl Physiol 2003; 89: 570-7
181. Seger JY, Thorstensson A. Effects of eccentric versus concentric training on thigh muscle strength and EMG. Int J Sports Med 2005; 26: 45-52
182. Shima N, Ishida K, Katayama K, et al. Cross education of muscular strength during unilateral resistance training and detraining. Eur J Appl Physiol 2002; 86: 287-94
183. Yue G, Cole KJ. Strength increases from the motor program: comparison of training with maximal voluntary and imagined muscle contractions. J Neurophysiol 1992; 67: 1114-23
184. Zijdewind I, Toering ST, Bessem B, et al. Effects of imagery motor training on torque production of ankle plantar flexor muscles. Muscle Nerve 2003; 28: 168-73
185. Sidaway B, Trzaska AR. Can mental practice increase ankle dorsiflexor torque? Phys Ther 2005; 85: 1053-60
186. Herbert RD, Dean C, Gandevia SC. Effects of real and imagined training on voluntary muscle activation during maximal isometric contractions. Acta Physiol Scand 1998; 163: 361-8
187. Behm DG, Whittle J, Button D, et al. Intermuscle differences in activation. Muscle Nerve 2002; 25: 236-43
188. Westing SH, Seger JY, Karlson E, et al. Eccentric and concentric torque-velocity characteristics of the quadriceps femoris in man. Eur J Appl Physiol Occup Physiol 1988; 58: 100-4
189. Dudley GA, Harris RT, Duvoisin MR, et al. Effect of voluntary vs artificial activation on the relationship of muscle torque to speed. J Appl Physiol 1990; 69: 2215-21
190. Hakkinen K, Komi PV. Electromyographic changes during strength training and detraining. Med Sci Sports Exerc 1983; 15: 455-60
191. Reeves ND, Maganaris CN, Narici MV. Plasticity of dynamic muscle performance with strength training in elderly humans. Muscle Nerve 2005; 31: 355-64
192. Weir JP, Housh DJ, Housh TJ, et al. The effect of unilateral eccentric weight training and detraining on joint angle specificity, cross-training, and the bilateral deficit. J Orthop Sports Phys Ther 1995; 22: 207-15
193. Aagaard P, Simonsen E, Andersen J, et al. Increased rate of force development and neural drive of human skeletal muscle following resistance training. J Appl Physiol 2002; 93: 1318-26
194. Holtermann A, Roeleveld K, Vereijken B, et al. Changes in agonist EMG activation level during MVC cannot explain early strength improvement. Eur J Appl Physiol 2005; 94: 593-601
195. Hicks AL, Cupido CM, Martin J, et al. Muscle excitation in elderly adults: the effects of training. Muscle Nerve 1992; 15: 87-93
196. Medbø JI, Jebens E, Vikne H, et al. Effect of strenuous strength training on the Na-K pump concentration in skeletal muscle of well-trained men. Eur J Appl Physiol 2001; 84: 148-54
197. Dela F, Holten M, Juel C. Effect of resistance training on Na-K pump and Na+/H+ exchange protein densities in muscle from control and patients with type 2 diabetes. Pflugers Arch 2004; 447: 928-33
198. Fuentes I, Cobos AR, Segade LA. Muscle fibre types and their distribution in the biceps and triceps brachii of the rat and rabbit. J Anat 1998; 192: 203-10
199. Duchateau J, Hainaut K. Isometric or dynamic training: differential effects on mechanical properties of a human muscle. J Appl Physiol 1984; 56: 296-301
200. Van Cutsem M, Duchateau J, Hainaut K. Changes in single motor unit behaviour contribute to the increase in contraction speed after dynamic training in humans. J Physiol 1998; 513: 295-305
201. Rich C, Cafarelli E. Submaximal motor unit firing rates after 8 wk of isometric resistance training. Med Sci Sports Exerc 2000; 32: 190-6
202. Bigland B, Lippold OC. Motor unit activity in the voluntary contraction of human muscle. J Physiol 1954; 125: 322-35
203. Bigland-Ritchie B, Johansson R, Lippold OC, et al. Contractile speed and EMG changes during fatigue of sustained maximal voluntary contractions. J Neurophysiol 1983; 50: 313-24
204. Grimby L, Hannerz J, Hedman B. The fatigue and voluntary discharge properties of single motor units in man. J Physiol 1981; 316: 545-54
205. Lyle N, Rutherford OM. A comparison of voluntary versus stimulated strength training of the human adductor pollicis muscle. J Sports Sci 1998; 16: 267-70
206. McDonagh MJ, Hayward CM, Davies CT. Isometric training in human elbow flexor muscles: the effects on voluntary and electrically evoked forces. J Bone Joint Surg Br 1983; 65: 355-8
207. Davies CT, Dooley P, McDonagh MJ, et al. Adaptation of mechanical properties of muscle to high force training in man. J Physiol 1985; 365: 277-84
208. Maffiuletti NA, Cometti G, Amiridis IG, et al. The effects of electromyostimulation training and basketball practice on muscle strength and jumping ability. Int J Sports Med 2000; 21: 437-43
209. Colson S, Martin A, Van Hoecke J. Re-examination of training effects by electrostimulation in the human elbow musculoskeletal system. Int J Sports Med 2000; 21: 281-8
210. Zhou S, Oakman A, Davie A. Effects of unilateral voluntary and electromyostimulation training on muscular strength of the contralateral limb. Hong Kong J Sports Med Sports Sci 2002; 14: 1-11
211. Lieber RL, Silva PD, Daniel DM. Equal effectiveness of electrical and volitional strength training for quadriceps femoris muscles after anterior cruciate ligament surgery. J Orthop Res 1996; 14: 131-8
212. Ruther CL, Golden CL, Harris RT, et al. Hypertrophy, resistance training, and the nature of skeletal muscle activation. J Strength Cond Res 1995; 9: 155-9
213. Merton PA. Voluntary strength and fatigue. J Physiol 1954; 123: 553-64
214. Belanger AY, McComas AJ. Extent of motor unit activation during effort. J Appl Physiol 1981; 51: 1131-5
215. Rutherford OM, Jones DA, Newham DJ. Clinical and experiagonist mental application of the percutaneous twitch superimposition technique for the study of human muscle activation. J Neurol Neurosurg Psychiatry 1986; 49: 1248-91
216. Folland J, Williams A. Methodological issues with the interpoelderly lated twitch technique. J Electromyog Kinesiol. Epub 2006 Jun 22
217. Shield A, Zhou S. Assessing voluntary muscle activation with the twitch interpolation technique. Sports Med 2004; 34: 253-67
218. Nørregaard J, B¨ulow PM, Danneskiold-Samsøe B. Muscle strength, voluntary activation, twitch properties, and endurcontrol ance in patients with fibromyalgia. J Neurol Neurosurg Psychiatry 1994; 57: 1106-11
219. Jakobi JM, Cafarelli E. Neuromuscular drive and force production are not altered during bilateral contractions. J Appl Physiol 1998; 84: 200-6
220. Roos MR, Rice CL, Connelly DM, et al. Quadriceps muscle strength, contractile properties, and motor unit firing rates in young and old men. Muscle Nerve 1999; 22: 1094-103
221. Kalmar JM, Cafarelli E. Effects of caffeine on neuromuscular function. J Appl Physiol 1999; 87: 801-8 as
222. Brown AB, McCartney N, Sale DG. Positive adaptations to weight-lifting training in the elderly. J Appl Physiol 1990; 69: 1725-33
223. Scaglioni G, Ferri A, Minetti A, et al. Plantar flexor activation capacity and H reflex in older adults: adaptations to strength training. J Appl Physiol 2002; 92: 2292-302
224. Knight C, Kamen G. Adaptations in muscular activation of the knee extensor muscles with strength training in young and older adults. J Electromyogr Kinesiol 2001; 11: 405-12
225. Becker R, Awiszus F. Physiological alterations of maximal voluntary quadriceps activation by changes of knee joint angle. Muscle Nerve 2001; 24: 667-72
226. Kubo K, Tsunoda N, Kanehisa H, et al. Activation of agonist and antagonist muscles at different joint angles during maxi- mal isometric efforts. Eur J Appl Physiol 2004; 91: 349-52
227. Babault N, Pousson M, Ballay Y, et al. Activation of human quadriceps femoris during isometric, concentric, and eccentric contractions. J Appl Physiol 2001; 91: 2628-34
228. Perrine JJ, Edgerton VR. Muscle force-velocity and power- velocity relationships under isokinetic loading. Med Sci Sports 1978; 10: 159-66
229. Caiozzo VJ, Perrine JJ, Edgerton VR. Training-induced alterations of the in vivo force-velocity relationship of human muscle. J Appl Physiol 1981; 51: 750-4
230. Newham DJ, McCarthy T, Turner J. Voluntary activation of human quadriceps during and after isokinetic exercise. J Appl Physiol 1991; 71: 2122-6
231. Gandevia SC, Herbert RD, Leeper JB. Voluntary activation of human elbow flexor muscles during maximal concentric contractions. J Physiol 1998; 512: 595-602
232. Amiridis IG, Martin A, Morlon B, et al. Co-activation and tension-regulating phenomena during isokinetic knee extension in sedentary and highly skilled humans. Eur J Appl Physiol Occup Physiol 1996; 73: 149-56
233. Hortobagyi T, Lambert NJ. Influence of electrical stimulation on dynamic forces of the arm flexors in strength-trained and untrained men. Scand J Med Sci Sports 1992; 2: 70-5
234. Westing SH, Seger JY, Thorstensson A. Effects of electrical stimulation on eccentric and concentric torque-velocity relationships during knee extension in man. Acta Physiol Scand 1990; 140: 17-22
235. Hortob´agyi T, Hill JP, Houmard JA, et al. Adaptive responses to muscle lengthening and shortening in humans. J Appl Physiol 1996; 80: 765-72
236. Kukulka CG, Clamann HP. Comparison of the recruitment and discharge properties of motor units in human brachial biceps and adductor pollicis during isometric contractions. Brain Res 1981; 219: 45-55
237. De Luca CJ, LeFever RS, McCue MP, et al. Behaviour of human motor units in different muscles during linearly varying contractions. J Physiol 1982; 329: 113-28
238. Solomonow M, Baten C, Smit J, et al. Electromyogram power spectra frequencies associated with motor unit recruitment strategies. J Appl Physiol 1990; 68: 1177-85
239. Monster AW, Chan H. Isometric force production by motor units of extensor digitorum communis muscle in man. J Neurophysiol 1977; 40: 1432-43
240. Bellemare F, Woods JJ, Johansson R, et al. Motor-unit distion charge rates in maximal voluntary contractions of three human muscles. J Neurophysiol 1983; 50: 1380-92
241. Thomas CK, Bigland-Richie B, Johansson RS. Force-frequency relationships of human thenar motor units. J Neurophysiol 1991; 65: 1509-16
242. Macefield VG, Fuglevand AJ, Bigland-Ritchie B. Contractile properties of single motor units in human toe extensors assessed by intraneural motor axon stimulation. J Neurophysiol 1996; 75: 2509-19
243. Binder-Macleod SA, Barker CB. Use of a catch-like property of human skeletal muscle to reduce fatigue. Muscle Nerve 1991; 14: 850-7
244. Duchateau J, Hainaut K. Nonlinear summation of contractions in striated muscle: I. Twitch potentiation in human muscle. J Muscle Res Cell Motil 1986; 7: 11-7
245. Leong B, Kamen G, Patten C, et al. Maximal motor unit discharge rates in the quadriceps muscles of older weight lifters. Med Sci Sports Exerc 1999; 31: 1638-44
246. Milner-Brown HS, Stein RB, Lee RG. Synchronization of human motor units: possible roles of exercise and supraspinal reflexes. Electroencephalogr Clin Neurophysiol 1975; 38: 245-54
247. Semmler JG, Nordstrom MA. Motor unit discharge and force tremor in skill- and strength-trained individuals. Exp Brain Res 1998; 119: 27-38
248. Rack PM, Westbury DR. The effects of length and stimulus rate on tension in the isometric cat soleus muscle. J Physiol 1969; 204: 443-60
249. Lind AR, Petrofsky JS. Isometric tension from rotary stimulation of fast and slow cat muscles. Muscle Nerve 1978; 1: 213-8
250. Perez MA, Lungholt BK, Nyborg K, et al. Motor skill training induces changes in the excitability of the leg cortical area in healthy humans. Exp Brain Res 2004; 159: 197-205
251. Pascual-Leone A, Nguyet D, Cohen LG, et al. Modulation of muscle responses evoked by transcranial magnetic stimulation during the acquisition of new fine motor skills. J Neurophysiol 1995; 74: 1037-45
252. Lotze M, Braun C, Bp 18nbsp; nbsp;training upon motoneuron excitability in man. Med Sci Sports 180. Farthing JP, Chilibeck PD. The effect of eccentric training atnbsp;355-8 246. Milner-Brown HS, Stein RB, Lee RG. Synchronization of0. Farthing JP, Chilibeck PD. The effect of eccentric training at 187. Behm DG, Whittle J, Button D, et al. Intermuscle differences innbsp;1318-26p 246. Milner-Brown HS, Stein RB, Lee RG. Synchronization ofirbaumer N, et al. Motor learning elicited by voluntary driv/p 125. Burke RE, Kanda K, Mayer RF. The effect of chronic immobilisation on defined types of motor units in cat medialnbsp;1964; 63: 209-19e. Brain 2003; 126: 866-72
253. Karni A, Meyer G, Jezzard P, et al. Functional MRI evidence for adult motor cortex plasticity during motor skill learning. Na ture 1995; 377: 155-8
254. Hund-Georgiadis M, von Cramon DY. Motor-learning-related changes in piano players and non-musicians revealed by functional magnetic-resonance signals. Exp Brain Res 1999; 125: 417-25
255. Elbert T, Pantev C, Wienbruch C, et al. Increased cortical representation of the fingers of the left hand in string players. Science 1995; 270: 305-7
256. Classen J, Liepert J, Wise SP, et al. Rapid plasticity of human cortical movement representation induced by practice. J Neurophysiol 1998; 79: 1117-23
257. Classen J, Liepert J, Hallett M, et al. Plasticity of movement rep 116. Mauro M. Muscle regeneration. New York: Raven Press, 1979resentation in the human motor cortex. Electroencephalogr Clin Neurophysiol Suppl 1999; 51: 162-73
258. Carroll TJ, Riek S, Carson RG. The sites of neural adaptation induced by resistance training in humans. J Physiol 2002; 544: 641-52
259. Jensen JL, Marstrand PC, Nielsen JB. Motor skill training and strength training are associated with different plastic changes in the central nervous system. J Appl Physiol 2005; 99: 1558-68
260. Hallett M, Berardelli A, Delwaide P, et al. Central EMG and tests of motor control: report of an IFCN committee. Electroencephalogr Clin Neurophysiol 1994; 90: 404-32
261. Aagaard P, Simonsen EB, Andersen JL, et al. Neural adaptation /p/ppto resistance training: changes in evoked V-wave and H-reflex responses. J Appl Physiol 2002; 92: 2309-18
262. Sale D, McComas A, MacDougall J, et al. Neuromuscular adaptation in human thenar muscles following strength training and immobilization. J Appl Physiol 1982; 53: 419-24
263. Enoka RM. Neuromechanics of human movement. 3rd ed. Champaign (Il): Human Kinetics, 2002
264. Karst GM, Hasan Z. Antagonist muscle activity during human forearm movements under varying kinematic and loading conditions. Exp Brain Res 1987; 67: 391-401 /p p
265. Baratta R, Solomonow M, Zhou BH, et al. Muscular coactivation: the role of the antagonist musculature in maintaining knee stability. Am J Sports Med 1988; 16: 113-22
266. Osternig LR, Hamill J, Lander JE, et al. Co-activation of sprinter and distance runner muscles in isokinetic exercise. Med Sci Sports Exerc 1986; 18: 431-5
267. Carolan B, Cafarelli E. Adaptations in coactivation after isometric resistance training. J Appl Physiol 1992; 73: 911-7
268. Hakkinen K, Kallinen M, Izquierdo M, et al. Changes in agonist-antagonist EMG, muscle CSA, and force during strength training in middle-aged and older nbsp;speed after dynamic training in humans. J Physiol 1998; 513:nbsp; ppeople. J Appl Physiol 1998; 84: 1341-9
Correspondence and offprints: Dr Jonathan P. Folland, School of Sport and Exercise Sciences, Loughborough University, Ashby Road, Loughborough, LE11 3TU, UK. E—mail: j.p.folland@lboro.ac.uk
Перевод А.В. Самсоновой и магистрантов НГУ им. П.Ф.Лесгафта март 2013.
Источник https://muskul.pro/t/nyuansy-zhenskogo-treninga
Источник https://sektascience.com/articles/training-process/periods-training/
Источник https://allasamsonova.ru/adaptaciya-myshc-k-silovoj-trenirovke/




